The science of the stomata of plants: a continuously growing list of references, abstracts and illustrations, helping researchers to data on publications.
Figure 2 IDD16-OE plants have lower stomatal densities. (a) The midleaf region of the eighth rosette leaf was used for analysing the stomatal phenotype. (b–d) Images of abaxial epidermal layers of the eighth leaf from 5-week-old wild-type (Col-0) and IDD16-OE plants. (e–h) Stomata density, pavement cell density, the number of stomatal precursor cells, and stomatal index were analysed in the eighth leaf abaxial epidermal layers from wild-type and IDD16-OE plants. Three areas in the midleaf region were imaged per leaf. Data are the mean of six individual plants (mean SE, n = 6). Asterisks indicate statistical significance based on Student’s t test. ** P < 0.01; * P < 0.05. (i–q) Confocal images of the cotyledon epidermis of wild-type, 35S:IDD16 m and 35S: IDD16s seedlings between 2 and 5 DAG. Cell outlines were visualized by propidium iodide (red) staining and the white arrows indicate the precursor cells. Scale bars: 50 lm.
IDD16 negatively regulates stomatal initiation via trans-repression of SPCH in Arabidopsis
by Qi S.-L., Lin Q.-F., Feng X.-J., Han H.-L., Liu J., Zhang L., Wu S., Le J., Blumwald E., Hua X.-J. (2019)
Shi-Lian Qi1,3,7,† , Qing-Fang Lin1,4,7,†, Xuan-Jun Feng1,7, Hui-Ling Han1,7, Jie Liu1,7, Liu Zhang6, Shuang Wu3, Jie Le5,7,* Eduardo Blumwald8 and Xue-Jun Hua1,2,*
1 Key Laboratory of Plant Resources and Beijing Botanical Garden, Institute of Botany, Chinese Academy of Sciences, Beijing, China 2 College of Life Sciences, Zhejiang Sci-Tech University, Hangzhou, Zhejiang, China 3 College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou, Fujian, China 4 College of Crop Science, Fujian Agriculture and Forestry University, Fuzhou, Fujian, China 5 Key Laboratory of Plant Molecular Physiology, CAS Center for Excellence in Molecular Plant Sciences, Institute of Botany, Chinese Academy of Sciences, Beijing, China 6 College of Life Sciences, Fujian Agriculture and Forestry University, Fuzhou, Fujian, China 7 University of Chinese Academy of Sciences, Beijing, China 8 Department of Plant Sciences, University of California, Davis, CA, USA
===
In Plant Biotechnology Journal 17: 1446–1457 – doi: 10.1111/pbi.13070 –
Figure 3 Effect of IDD16 overexpression on stomatal development in the leaf adaxial epidermis. (a–c) Images of leaf adaxial epidermal layers of the eighth leaf from 5-week-old wildtype (Col-0) and IDD16-OE plants. (d–f) Stomatal density, stomatal index and pavement cell density were analysed in the leaf adaxial epidermis of the eighth leaf from wild-type and IDD16-OE plants. Three areas in the midleaf region were imaged per leaf. Data are the mean of six individual plants (mean SE, n = 6). Asterisks indicate statistical significance based on Student’s t test. ** P < 0.01; * P < 0.05. (g–o) Confocal images of the cotyledon adaxial epidermis of wild-type (Col0), 35S:IDD16m and 35S:IDD16s seedlings between 2, 3, 5 DAG. Cell outlines were visualized by PI staining and the white arrows indicate the precursor cells. Scale bars: 50 lm.
Summary
In Arabidopsis, the initiation and proliferation of stomatal lineage cells is controlled by SPEECHLESS (SPCH). Phosphorylation of SPCH at the post-translational level has been reported to regulate stomatal development. Here we report that IDD16 acts as a negative regulator for stomatal initiation by directly regulating SPCH transcription. In Arabidopsis,
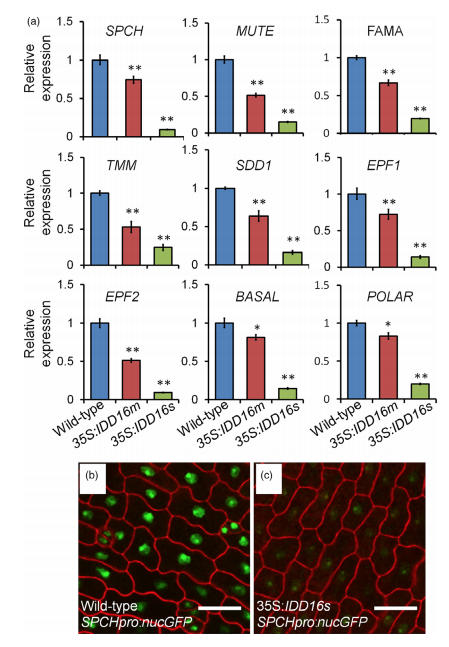
Figure 4 IDD16 inhibits stomatal initiation by negatively regulating SPCH expression. (a) Analysis of the expression of genes related to stomatal development. qRT-PCR was performed on total RNA from 4-DAG wild-type and IDD16-OE plants. The expression level in wild-type was normalized as 1. Values are means SE from three biological replicates. Asterisks indicate statistical significance based on Student’s t test; ** P < 0.01; * P < 0.05. (b) and (c) Confocal images of SPCHpro:nucGFP abaxial cotyledons in wild-type (b) and 35S: IDD16s (c) background, respectively. Epidermal cell periphery is highlighted by propidium iodide (red) staining. Scale bars: 25 lm.
IDD16 overexpression decreased abaxial stomatal density in a dose-dependent manner. Time course analysis revealed that the initiation of stomatal precursor cells in the IDD16-OE plants was severely inhibited. Consistent with these findings, the transcription of SPCH was greatly repressed in the IDD16-OE plants. In contrast, IDD16-RNAi transgenic line resulted in enhanced stomatal density, suggesting that IDD16 is an intrinsic regulator of stomatal development. ChIP analysis indicated that IDD16 could directly bind to the SPCH promoter.
Figure 5 IDD16 negatively regulates stomatal development. (a) Confocal microscopy image of IDD16pro:IDD16-GFP showing nuclear localization of IDD16 in adaxial epidermis of cotyledons in 1-DAG seedlings. Epidermal cell periphery is visualized by propidium iodide (red) staining. White circles indicate meristemoids (Ms). (b and c) Compared to wild-type (b), IDD16pro:IDD16-GFP transgenic line (c) showed reduced stomatal density in abaxial epidermis of the cotyledons from 8-DAG seedlings. Scale bars: 50 lm. (d) The expression levels of IDD16 in 4-DAG wild-type and IDD16-RNAi seedlings were determined by qRT-PCR. Values are means SE from three biological replicates. (e–g) Compared to wild-type Col (e), IDD16-RNAi plants (f, g) showed increased stomatal density in abaxial epidermis of the cotyledons from 8-DAG seedlings. Scale bars: 50 lm. (h) Stomata density obtained from the samples in (e–g). Three areas in the midleaf region were imaged per leaf. Data are the mean of six individual plants (mean SE, n = 6). Asterisks indicate statistical significance based on Student’s t test; ** P < 0.01.
Furthermore, Arabidopsis plants overexpressing IDD16 exhibited significantly increased drought tolerance and higher integrated water use efficiency (WUE) due to reduction in leaf transpiration. Collectively, our results established that IDD16 negatively regulates stomatal initiation via trans-repression of SPCH, and thus provide a practical tool for increasing plant WUE through the manipulation of IDD16 expression.
Organic electronic ion pump delivering plant hormones to leaf tissue for stomata control – Electronic plants designed to be more resilient to environmental stress could offer major advantages in agriculture. IMAGE CREDIT: THOR BALKHED
Bioelectronic Implants for Drought-Resistant Plants
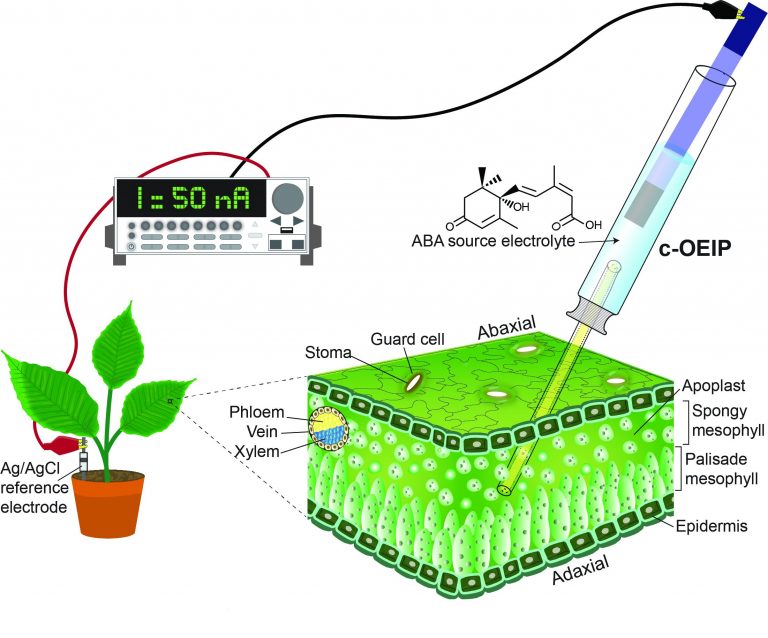
Delivery of ABA into the spongy mesophyll layer of a plant.
The merging of electronics with biology—bioelectronics—is a rapidly growing field of interest in materials science and biomedical engineering. So far, research has mainly focused on healthcare applications, in which minimally invasive electronic devices that integrate with the human body in order to perform a specific function—monitoring a physiological process, diagnosing a disease, or repairing damaged tissue—have led to new ways of addressing a range of medical issues.
For example, biomedical implants—some of which can even be delivered by ingestion—that are designed to deteriorate after their use has been fulfilled is one fascinating technology to emerge from bioelectronics research. Not only does this ensure that the device remains in the body only for its period of clinical utility but renders risky surgical procedures to remove it unnecessary.
The possibility of extending bioelectronics to other organisms—plants—is also beginning to be explored. Engineering plants with a higher tolerance to environmental stress caused by extreme weather conditions such as drought is becoming especially important as the effects of the changing climate manifest.
Dr. Eleni Stavrinidou, who leads the Electronic Plants (e-Plants) subgroup in the Laboratory of Organic Electronics at Linköping University, Sweden, is one researcher pioneering this field. In a recent paper in Small, she presents a device designed to deliver the phytohormone abscisic acid (ABA) into an intact tobacco plant—the first example of an implantable bioelectronic device in plants.
ABA, a large biomolecule, is one of the main hormones involved in regulating plant response to stress, inducing closure of the stomata in drought conditions in order to restrict water loss. Stomata also play a key role in photosynthesis by regulating gas exchange.
The organic electronic ion pump (c-OEIP) developed by her team is minimally invasive for the plant and delivers ABA electronically into the leaf apoplast, inducing closure of the stomata. Conventional OEIP devices have large dimensions and use planar geometries that hinder implantation into soft tissue, whereas this novel, miniaturized device based on glass capillary fibers has both considerably reduced device dimensions and a simplified fabrication process.
“This work paves the way for establishing bioelectronic devices as versatile tools for biology and the use of bioelectronic devices not only in biomedicine but also in agriculture and forestry,” states Dr. Stavrinidou.
Achieving controlled delivery of even larger signaling molecules, with more dynamic capacity and single-cell resolution, is the next challenge she anticipates in this research.
Stomata are the structural elements of plant epidermis which control transpiration and gas exchange. Each stoma consists of two guard cells divided by the stomatal aperture. These cells are capable of reversible deformations determining the width of aperture. It is known that these deformations depend on the value of turgor pressure in the guard cells and on the structure of their walls. In this work, the influence of the outer tangential wall geometry of the guard cells on stomatal movements is estimated by means of the finite element method in the ANSYS software. The application of modelling has shown that cuticular outgrowths on the tangential walls influence the degree and pattern of guard cell deformations. The outgrowths prevent wide opening of the stomatal aperture and cause its sinking deep into leaf epidermis. The functional significance of such stomatal movements is discussed. It is deduced that the discovered phenomenon had great importance to the survival of laurophyllous plants in conditions of aridization.
Increasing vapor pressure deficit (VPD) increases atmospheric demand for water, and vapor pressure deficit is expected to rise with increasing greenhouse gases. While increased evapotranspiration (ET) in response to increased atmospheric demand seems intuitive, plants are capable of reducing ET in response to increased VPD by closing their stomata, in an effort to conserve water. Here we examine which effect dominates response to increasing VPD: atmospheric demand and increases in ET, or plant physiological response (stomata closure) and decreases in ET. We use Penman-Monteith, combined with semi-empirical optimal stomatal regulation theory and underlying water use efficiency, to develop a theoretical framework for understanding how ET responds to increases in VPD.
The theory suggests that for most environmental conditions and plant types, plant physiological response dominates and ET decreases with increasing VPD. Plants that are evolved or bred to prioritize primary production over water conservation (e.g. crops) exhibit a higher likelihood of atmospheric demand-driven response (ET increasing). However for forest, grass, savannah, and shrub plant types, ET more frequently decreases than increases with rising VPD. This work serves as an example of the utility of our simplified framework for disentangling land-atmosphere feedbacks, including the characterization of ET response in an atmospherically drier, enriched CO2 world.
Nine conifer species with narrow (<5 mm), single-veined leaves were selected for the purpose of examining changes in intercellular CO2 concentration (ci) during drought. Due to the leaf morphology of the study plants, the confounding effects of nonhomogenous photosynthesis common to most reticulate-veined angiosperms were largely avoided, giving a clear picture of ci dynamics under increasing drought. A characteristic biphasic response was observed in all species, with an initial stomatal control phase resulting in a substantial reduction in ci as stomatal conductance (gs) decreased. As gs reached low levels, a strong nonstomatal limitation phase was observed, causing ci to increase as gs approached a minimum. This nonstomatal phase was linked to a concomitant rapid decrease in the fluorescence parameter quantum efficiency, indicating the onset of nonreversible photoinhibition. The ratio of internal to atmospheric CO2 concentration (ci/ca) decreased from values of between 0.68 and 0.57 in undroughted plants to a minimum, (ci/ca)min, which was well defined in each species, ranging from 0.10 in Actinostrobus acuminatus to 0.36 in Acmopyle pancheri. A high correlation was found to exist between (ci/ca)min and leaf water potential measured at (ci/ca)min. Species developing high maximum intrinsic water use efficiencies (low [ci/ca]min), such as A. acuminatus, did so at lower leaf water potentials (-4.5 MPa) than more mesic species (-1.75 MPa for A. pancheri). It is concluded that in the absence of patchy stomatal closure, (ci/ca)min gives a good representation of the drought tolerance of foliage.
Definitions of the variables used and the units are given in Table 1
The literature reports enormous variation between species in the extent of stomatal responses to rising CO2. This paper attempts to provide a framework within which some of this diversity can be explained. We describe the role of stomata in the short‐term response of leaf gas exchange to increases in ambient CO2 concentration by developing the recently proposed stomatal model of 26). In this model stomatal conductance is correlated with the functioning of the photosynthetic system so that the effects of increases in CO2 on stomata are experienced through changes in the rate of photosynthesis in a simple and mechanistically transparent way. This model also allows us to consider the effects of evaporative demand and soil moisture availability on stomatal responses to photosynthesis and therefore provides a means of considering these additional sources of variation. We emphasize that the relationship between the rate of photosynthesis and the internal CO2 concentration and also drought will have important effects on the relative gains to be achieved under rising CO2.
Stomatal Response to Water Stress and its Relationship to Bulk Leaf Water Status and Osmotic Adjustment in Pearl Millet (Pennisetum americanum [L.] Leeke)
by Henson I. E., Alagarswamy G., Mahalakshmi V., Bidinger F. R., (1983)
In Journal of Experimental Botany 34(141): 442-450 –
The water potential (Ψ) at which stomata completed closure (Ψ8Lmin) was determined for pearl millet (Pennisetum americanum [L.] Leeke) at two growth stages by monitoring changes in leaf conductance (gL) and Ψ following shoot detachment. Leaf water status was evaluated concurrently using a pressure-volume (P-V) technique.
In a pot experiment with young vegetative plants, Ψ8Lmin closely approximated to the estimated Ψ at zero turgor (Ψu) both for control and for drought-conditioned plants which had osmotically adjusted. However, for penultimate leaves of field-grown flowering plants, Ψ8Lmin was found to be 0.61 (irrigated plants) and 0.87 (droughted plants) MPa below Ψu. In drought-stressed field-grown plants, osmotic adjustment (characterized by a decrease in solute (osmotic) potential (Ψs ) at both full hydration and zero turgor) was insufficient to maintain a positive bulk leaf turgor potential (Ψp) once Ψ had declined to below about -1.5 MPa. It is suggested that localized adjustment by the stomatal complex in response to environmental differences, leaf ageing and/or ontogenetic change, is responsible for the uncoupling of stomatal from bulk leaf water status.
Changes in the stomatal aperture and the rate of transpiration under foliar application with triazine (atrazine) urea (diuron), amide (alachlor, butachlor) and thiocarbamate (EPTC and Molinate) type of herbicides onAmaranthus viridis L.;Digera arvensis Forsk;Boerhaavia diffusa L.;B. repanda Willd.;Commelina benghalensis L.;Cyperus rotundus L.;Pisum sativum L.; andPennisetum typhoides Stapf and Hubb; were studied. It was observed that the triazine and urea herbicides have caused an appreciable stimulation of stomatal opening within two hours of chemical contact, while the amides possessed a contrasting behaviour in as much as they have brought a rapid closure of the open stomata. Thiocarbamates on the other hand have exhibited a differential response characteristically relating to the C3 or C4 type plants. The general trend observed with the thiocarbamates was that they were stimulatory for the C3 and inhibitory for the C4 plants in respect of stomatal opening.
A definite correlation of the rate of the transpiration with the stomatal response to the herbicide was found. The present study suggests that the resistance or susceptibility of the plants is determined by the selective stomatal behaviour towards different herbicides.
Implantable Bioelectronics: Implantable Organic Electronic Ion Pump Enables ABA Hormone Delivery for Control of Stomata in an Intact Tobacco Plant
by Bernacka‐Wojcik I., Huerta M., Tybrandt K., Karady M., Mulla M. Y., Poxson D. J., Gabrielsson E. O., Ljung K., Simon D. T., Berggren M., Stavrinidou E. (2019)
Iwona Bernacka‐Wojcik, Miriam Huerta, Klas Tybrandt, Michal Karady, Mohammad Yusuf Mulla, David J. Poxson, Erik O. Gabrielsson, Karin Ljung, Daniel T. Simon, Magnus Berggren, Eleni Stavrinidou,
The organic electronic ion pump (OEIP) provides a unique means for electronically controlled, flow‐free delivery of ions at the cellular scale. In article number 1902189, Eleni Stavrinidou and co‐workers implant a capillary‐based OEIP in a leaf of an intact tobacco plant with no significant wound response and effectively deliver the hormone abscisic acid, which is the first example of an implantable bioelectronic device in plants.
…………………………
Electronic control of biological processes with bioelectronic devices holds promise for sophisticated regulation of physiology, for gaining fundamental understanding of biological systems, providing new therapeutic solutions, and digitally mediating adaptations of organisms to external factors. The organic electronic ion pump (OEIP) provides a unique means for electronically‐controlled, flow‐free delivery of ions, and biomolecules at cellular scale. Here, a miniaturized OEIP device based on glass capillary fibers (c‐OEIP) is implanted in a biological organism. The capillary form factor at the sub‐100 µm scale of the device enables it to be implanted in soft tissue, while its hyperbranched polyelectrolyte channel and addressing protocol allows efficient delivery of a large aromatic molecule. In the first example of an implantable bioelectronic device in plants, the c‐OEIP readily penetrates the leaf of an intact tobacco plant with no significant wound response (evaluated up to 24 h) and effectively delivers the hormone abscisic acid (ABA) into the leaf apoplast. OEIP‐mediated delivery of ABA, the phytohormone that regulates plant’s tolerance to stress, induces closure of stomata, the microscopic pores in leaf’s epidermis that play a vital role in photosynthesis and transpiration. Efficient and localized ABA delivery reveals previously unreported kinetics of ABA‐induced signal propagation.
Stomata and the mesophyll must be developmentally coordinated on multiple scales. Individual stomata must overlie mesophyll airspaces, not cells, to form functional conduits for leaf gas exchange (a). For a given density of stomata, altering mesophyll architecture, for example by increasing the surface area of mesophyll cells exposed to airspaces (indicated in red), will alter the potential rates of gas flux, increasing the surface area available for CO2 uptake (b).
The developmental relationship between stomata and mesophyll airspace
Stomatal patterning and mesophyll development are not temporally separate processes. Distinct leaf layers are established very early in leaf development (a). Tiny airspaces at cell junctions arise in young leaves (indicated in red) concomitantly with stomatal patterning in the epidermis (b). The evidence suggests that while mesophyll airspace enlargement (c, indicated in red) is not entirely dependent upon the opening of stomatal pores, these processes are quantitatively correlated. Note: Arabidopsis leaves typically contain more layers of mesophyll cells than the two represented in these schematic diagrams.
Abstract
The quantitative and spatial coordination of stomatal pores in the epidermis and airspaces in the underlying mesophyll tissue is vital for efficient gas exchange in the leaf. Mechanisms that determine the distribution of stomata in the epidermis have been extensively studied, but how this relates to the regulation of mesophyll airspace configuration is poorly understood.
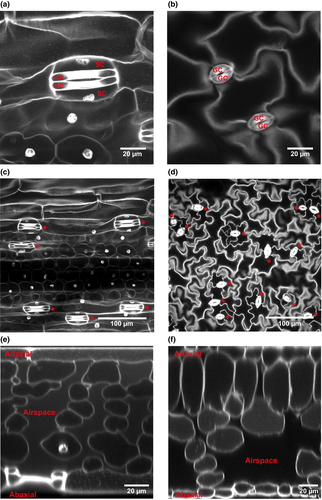
Monocots and eudicots differ in stomatal shape and patterning, and in their mesophyll structure. Monocot stomata have dumb‐bell‐shaped guard cells (GC) surrounded by subsidiary cells (SC) (a), whereas eudicot guard cells are kidney‐shaped and typically lack subsidiary cells (b). Stomata (*) in monocots are arranged in linear cell files (c) whereas the cells of the eudicot epidermis appear to be arranged more randomly (d). Monocot mesophyll is more porous towards the abaxial leaf surface (e) and this pattern is more pronounced in eudicots with clearly distinct adaxial palisade and abaxial spongy mesophyll layers (f). Confocal images of cleared and propidium iodide‐stained mature Arabidopsis (eudicot) and barley (monocot) leaves.
Recent studies have investigated how development is coordinated between these tissue layers. The evidence suggests that multiple mechanisms are likely to work concurrently to coordinate stomatal and mesophyll development for optimal leaf gas exchange, and that both genetic and physiological factors contribute to this regulation. Such advances in our understanding of leaf development have important implications for potential improvement of crop water use efficiency.
Summary of potential signals in stomata–mesophyll developmental coordination. The antagonistic peptides EPF1/EPF2 and STOMAGEN, derived from the epidermis and mesophyll respectively, compete for binding to the ER/TMM ligand receptor complex, which appears to be critical for coordination of stomatal and mesophyll development. In addition to these cell signals arising in relation to the development of stomata, stomatal function may also allow gas‐exchange‐based signals to play a role in setting the mesophyll structure.










You must be logged in to post a comment.