
Ecological Niches and Suitability Areas of Three Host Pine Species of Bark Beetle Dendroctonus mexicanus Hopkins

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Bioclimatic Variables and Selection
2.3. Species Occurrence Records and Cleaning
2.4. Calibration Area
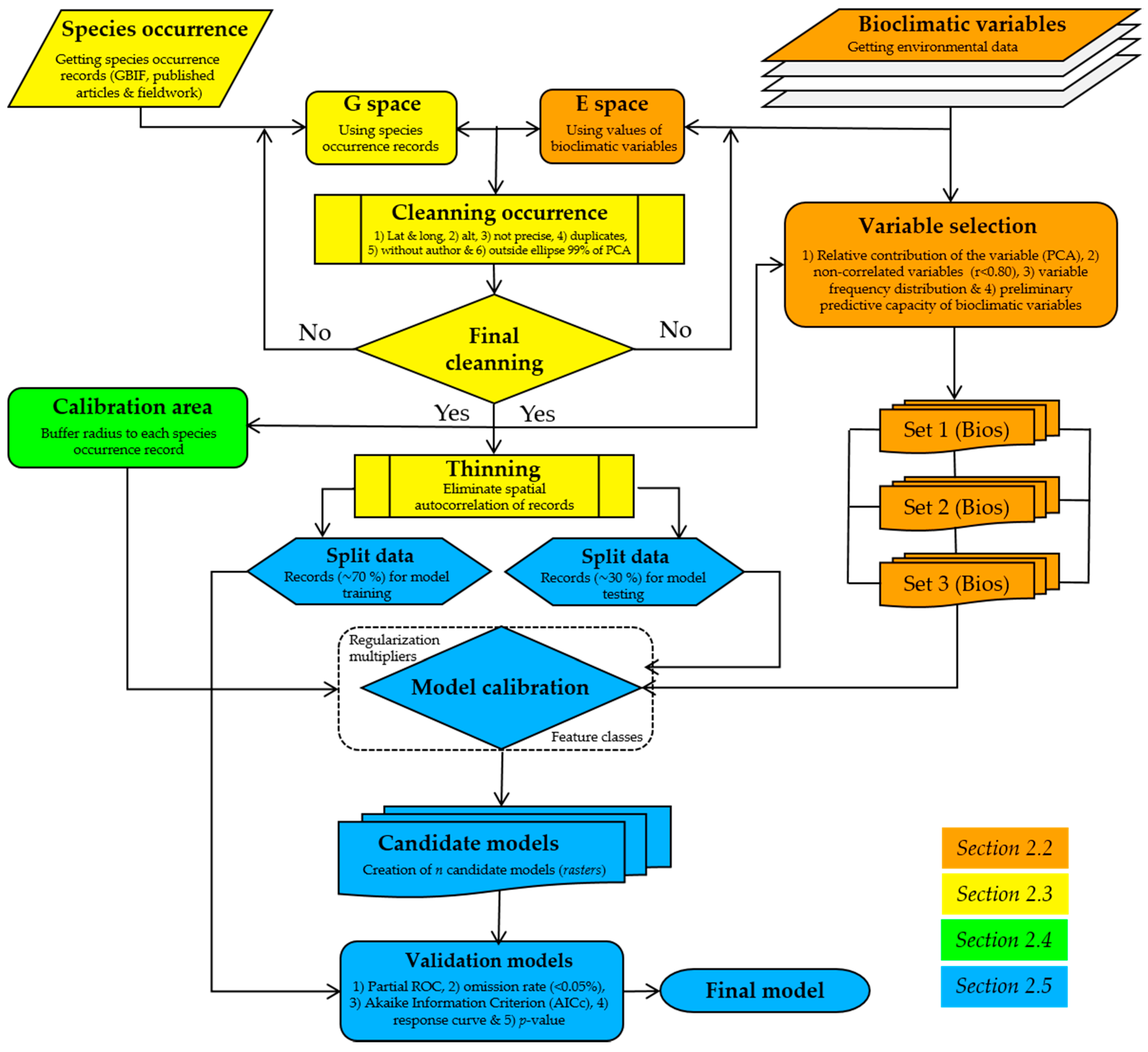
2.5. Model Calibration, Creation, and Evaluation
2.6. Model Stratification
2.7. Bark Beetle-Free Areas
2.8. Quantifying Niche Similarity
3. Results
3.1. Generalities
3.2. Generated Models and Their Statistics
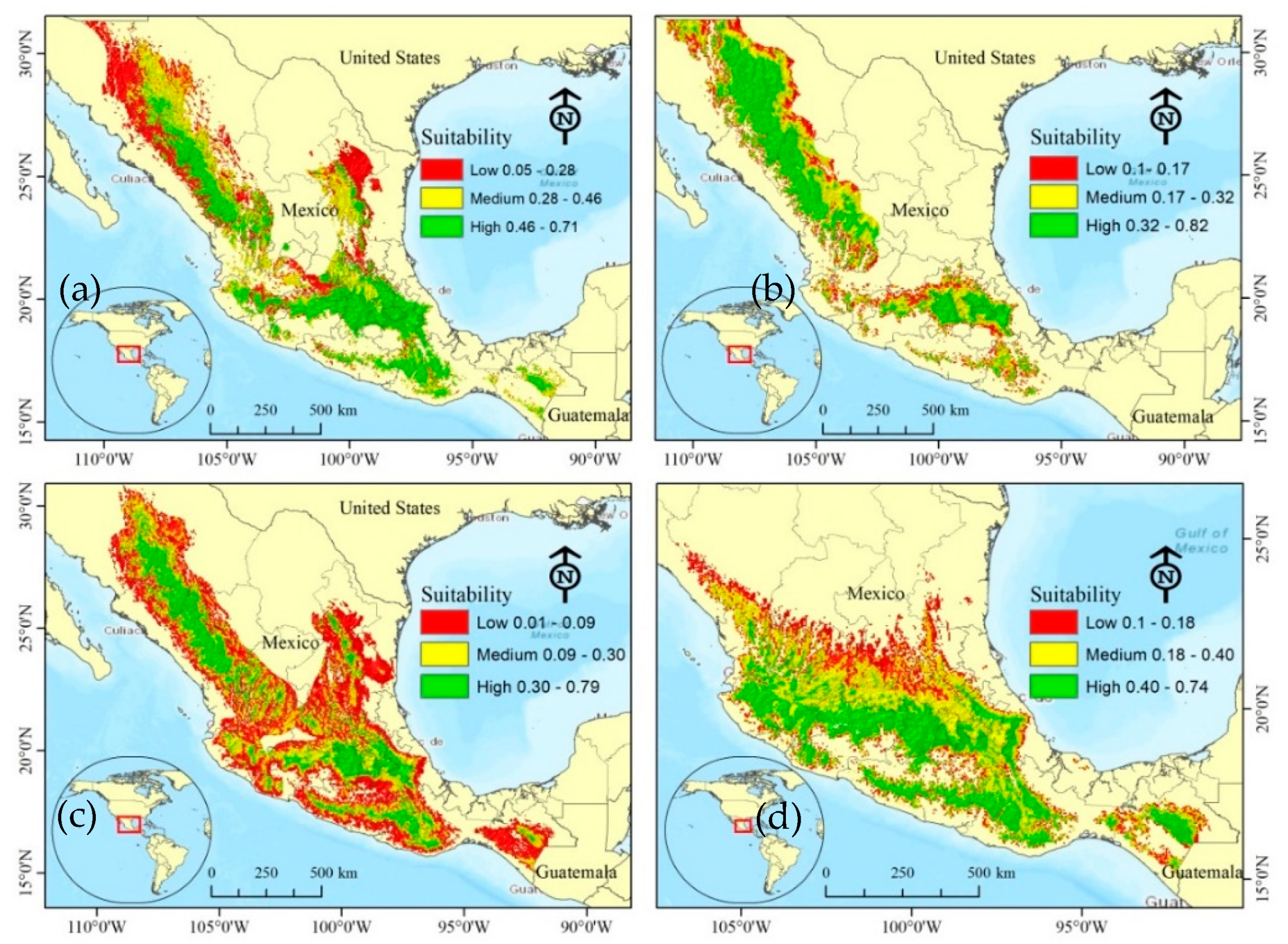
3.3. Species Suitability Areas
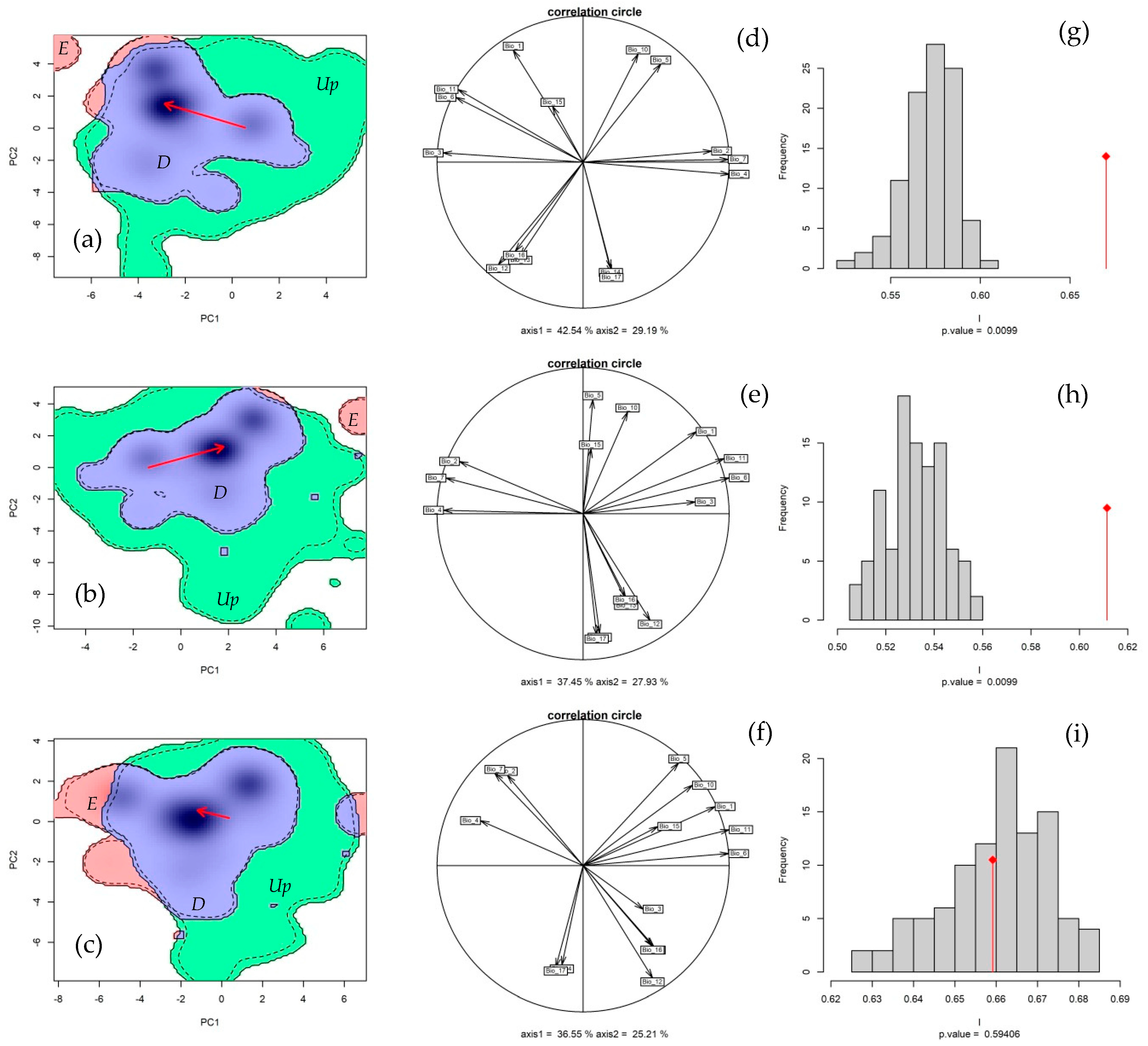
3.4. Bioclimatic Profile
3.5. Climatic Suitability of Pine Species, Free of Bark Beetle Suitability Areas
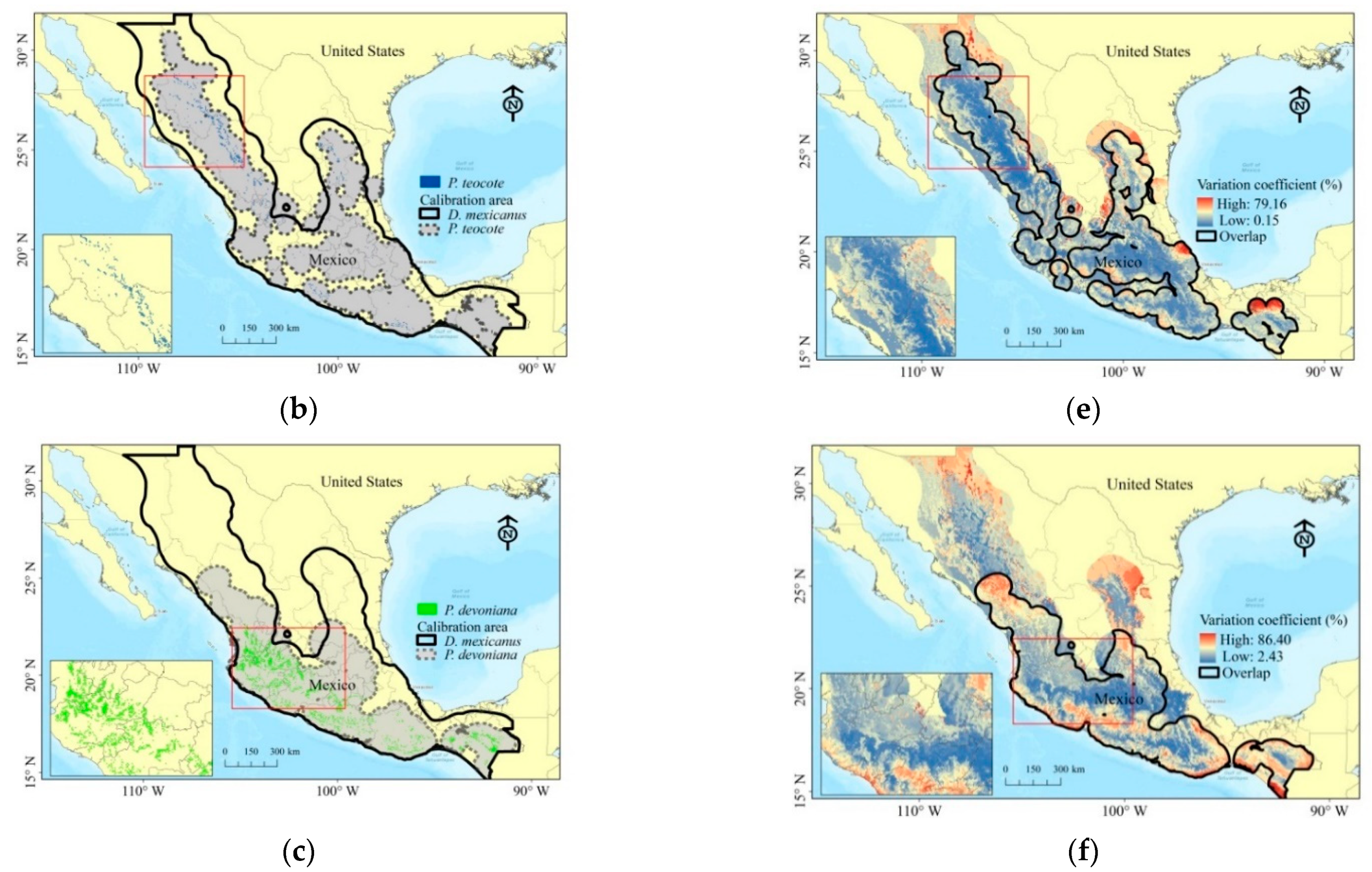
3.6. Overlap of Suitability and Ecological Niches
4. Discussion
4.1. Species Occurrence Records in ENM
4.2. Variable Importance in ENM
4.3. Climate Suitability and Niche Overlap
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Predictive Species Models
References
- del-Val, E.; Sáenz, R.C. Insectos Descortezadores (Coleoptera: Curculionidae) y Cambio Climático: Problemática Actual y Perspectivas En Los Bosques Templados. TIP Rev. Especializada en Cienc. Químico Biol. 2017, 20, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Kuennecke, B.H. Temperate Forest Biomes; Greenwood Press: London, UK, 2008; 194p. [Google Scholar]
- Challenger, A.; Soberón, J. Los ecosistemas terrestres. In Capital Natural de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Conabio, México, 2008. [Google Scholar]
- Rzedowsky, J. Vegetación de México, 1st ed.; Limusa: México City, México, 1978; p. 432. [Google Scholar]
- Gernandt, D.; Pérez-de la Rosa, J. Biodiversidad de Pinophyta (coníferas) en México. Rev. Mex. Biodivers. 2014, 85, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. El Estado de los Bosques del Mundo. las Vías Forestales Hacia el Desarrollo Sostenible; FAO: Roma, Italy, 2018; p. 52. [Google Scholar]
- Allen, M.R.; Dube, O.P.; Solecki, W.; Aragón-Durand, F.; Cramer, W.; Humphreys, S.; Kainuma, M.; Kala, J.; Mahowald, N.; Mulugetta, Y.; et al. Framing and Context. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; Masson-Delmotte, V., Zhai, V., Portner, H., Roberts, D., Skea, J., Shukla, P., Pirani, A., Moufouma-Okia, W., Chen, Y., Zhou, X., et al., Eds.; IPCC: Geneve, Switzerland, 2018. [Google Scholar]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae) a Taxonomic Monograph; Great Basin Naturalist Memoirs: Provo, UT, USA, 1982; p. 1359. [Google Scholar]
- Armendáriz, T.F.; Torres, B.V.; Fernanda, L.M.; Villa, C.J.; Zúñiga, G. New record and extension of the distribution range of the bark beetle Dendroctonus rhizophagus (Curculionidae: Scolytinae). Rev. Mex. Biodivers. 2012, 83, 850–853. [Google Scholar] [CrossRef]
- Bentz, B.J.; Jönsson, A.M. Modeling bark beetle responses to climate change. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 533–553. [Google Scholar]
- Six, D.; Bracewell, R. Dendroctonus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 305–350. [Google Scholar]
- Bentz, B.; Régnière, J.; Fettig, C.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.J.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Salinas, M.Y.; Ager, A.; Vargas, C.F.; Hayes, J.L.; Zúñiga, G. Determining the vulnerability of Mexican pine forests to bark beetles of the genus Dendroctonus Erichson (Coleoptera: Curculionidae: Scolytinae). For. Ecol. Manag. 2010, 260, 52–61. [Google Scholar] [CrossRef]
- Salinas, M.Y.; Mendoza, G.M.; Barrios, M.A.; Cisneros, R.; Macías, S.J.; Zúñiga, G. Areography of the genus Dendroctonus (Coleoptera: Cur-culionidae: Scolytinae) in México. J. Biogeogr. 2004, 31, 1163–1177. [Google Scholar] [CrossRef]
- Cibrián, T.D.; Méndez, M.J.; Campos, B.R.; Yates, H.O.; Flores, L.J. Insectos Forestales de México; Universidad Autónoma de Chapingo: Estado de México, México, 1995; p. 453. [Google Scholar]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez, M.E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Pricenton, NJ, USA, 2011. [Google Scholar]
- Cobos, M.E.; Jiménez, L.; Nuñez, P.C.; Romero, A.D.; Simões, M. Sample data and training modules for cleaning biodiversity information. Biodivers. Inform. 2018, 14, 49–50. [Google Scholar] [CrossRef] [Green Version]
- Méndez, E.F.; Méndez, G.J.; Cerano, P.J. Distribución actual y potencial de Dendroctonus mexicanus Hopkins bajo dos escenarios de cambio climático. Madera Y Bosques 2020, 26, 1–14. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Soberón, J.; Osorio, O.L.; Peterson, A.T. Diferencias conceptuales entre modelación de nichos y modelación de áreas de distribución. Rev. Mex. Biodivers. 2017, 88, 437–441. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Austin, M.P.; Van Niel, K.P. Improving species distribution models for climate change studies: Variable selection and scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Busby, J.R. A biogeoclimatic analysis of Nothofagus cunninghamii (Hook.) Oerst. In southeastern Australia. Aust. Ecol. 1986, 11, 1–7. [Google Scholar] [CrossRef]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- González, H.A.; Morales, V.R.; Romero, S.M.; Islas, T.B.; Pérez, M.R. Modelling potential distribution of a pine bark beetle in Mexican temperate forests using forecast data and spatial analysis tools. J. For. Res. 2020, 31, 649–659. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, T.A. Interpretation of models of fundamental ecological niches and species’ distributional areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harbor on Quantitative Symposia Biology. GS Search 1957, 22, 415–427. [Google Scholar]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Willis, K.J.; Whittaker, R.J. Species diversity-scale matters. Science 2002, 295, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria, A.R.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Escobar, L.E.; Lira, N.A.; Medina, V.G.; Peterson, A.T. Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: Use of Maxent and NicheA to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar] [CrossRef]
- Mendoza, M.G.; Salinas, M.Y.; Olivo, M.A.; Zúñiga, G. Factors influencing the geographical distribution of Dendroctonus rhizophagus (Coleoptera: Curculionidae: Scolytinae) in the Sierra Madre Occidental, Mexico. Environ. Entomol. 2011, 40, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- De Marco, P.; Nóbrega, C.C. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation. PLoS ONE 2018, 13, e0202403. [Google Scholar] [CrossRef]
- Sturges, H.A. The choice of a class interval. J. Am. Stat. Assoc. 1926, 21, 65–66. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 251–270. [Google Scholar] [CrossRef]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- Cruz, C.G.; López, M.L.; Villaseñor, J.L.; Ortiz, E. Potential species distribution modeling and the use of principal component analysis as predictor variables. Rev. Mex. Biodivers. 2014, 85, 189–199. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio, O.L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elith, J.; Kearney, M.; Phillips, S. The art of modelling range-shifting species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Morales, N.S.; Fernández, I.C.; Baca-González, V. MaxEnt’s parameter configuration and small samples: Are we paying attention to recommendations? A systematic review. PeerJ 2017, 5, e3093. [Google Scholar] [CrossRef]
- Smith, S.E.; Mendoza, M.G.; Zúñiga, G.; Halbrook, K.; Hayes, J.L.; Byrne, D.N. Predicting the distribution of a novel bark beetle and its pine hosts under future climate conditions. Agric. For. Entomol. 2013, 15, 212–226. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jimenez, V.A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2007, 17, 145–151. [Google Scholar] [CrossRef]
- Reddy, K.G.; Khan, M.G. stratify: An R Package for optimal stratification and sample allocation for univariate populations. Aust. N. Z. J. Stat. 2020, 62, 383–405. [Google Scholar] [CrossRef]
- Khan, E.A.; Khan, M.M.; Ahsan, M.J. Optimum stratification: A mathematical programming approach. Calcutta Stat. Assoc. Bull. 2002, 52, 323–333. [Google Scholar] [CrossRef]
- Khan, M.M.; Nand, N.; Ahmad, N. Determining the optimum strata boundary points using dynamic programming. Surv. Methodol. 2008, 34, 205–214. [Google Scholar]
- Khan, M.M.; Reddy, K.G.; Rao, D.K. Designing stratified sampling in economic and business surveys. J. Appl. Stat. 2015, 42, 2080–2099. [Google Scholar] [CrossRef]
- Schoener, T.W. Nonsynchronous spatial overlap of lizards in patchy habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 221, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Godsoe, W.; Case, B.S. Accounting for shifts in the frequency of suitable environments when testing for niche overlap. Methods Ecol. Evol. 2015, 6, 59–66. [Google Scholar] [CrossRef]
- Sánchez, G.A. Una visión actual de la diversidad y distribución de los pinos de México. Madera Y Bosques 2008, 14, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2016, 40, 774–787. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 11 May 2020).
- Loiselle, B.A.; Jørgensen, P.M.; Consiglio, T.; Jiménez, I.; Blake, J.G.; Lohmann, L.G.; Montiel, O.M. Predicting species distributions from herbarium collections: Does climate bias in collection sampling influence model outcomes. J. Biogeogr. 2008, 35, 105–116. [Google Scholar] [CrossRef]
- Shcheglovitova, M.; Anderson, R.P. Estimating optimal complexity for ecological niche models: A jackknife approach for species with small sample sizes. Ecol. Model. 2013, 269, 9–17. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning—ICML, Banff, AB, Canada, 4–8 July 2004. [Google Scholar]
- Austin, M. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Model. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Aguirre, G.J.; Serna, C.H.; Villalobos, A.A.; Pérez, D.J.; Raes, N. Similar but not equivalent: Ecological niche comparison across closely–related Mexican white pines. Divers. Distrib. 2015, 21, 245–257. [Google Scholar] [CrossRef]
- Williams, D.W.; Liebhold, A.M. Climate change and the outbreak ranges of two North American bark beetles. Agric. For. Entomol. 2002, 4, 87–99. [Google Scholar] [CrossRef]
- Stockwell, D. Improving ecological niche models by data mining large environmental datasets for surrogate models. Ecol. Model. 2006, 192, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest insects and climate change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, M.J.; Cera, J.I.; Mendoza, A.R.; Sáenz, L.A.; Torres, O.M.; Bravo, P.L.; Alatorre, C.L. Distribución potencial de Dendroctonus valens mediante modelos de máxima entropía: Estado de California, E.U. Rev. Latinoam. El Ambiente Y Las Cienc. 2015, 6, 194–198. [Google Scholar]
- Pearman, P.B.; Randin, C.F.; Broennimann, O.; Vittoz, P.; Knaap, W.O.; Engler, R.; Le-Lay, G.; Zimmermann, N.; Guisan, A. Prediction of plant species distributions across six millennia. Ecol. Lett. 2008, 11, 357–369. [Google Scholar] [CrossRef]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.T.; Guisan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Name | Description | Species | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Dendroctonus mexicanus | Pinus leiophylla | Pinus teocote | Pinus devoniana | ||||||
| PC1 (48.7) | PC2 (18.9) | PC1 (43.9) | PC2 (29.5) | PC1 (39.6) | PC2 (26.0) | PC1 (35.0) | PC2 (29.1) | ||
| Bio 1 | Annual Mean Temperature¶ (°C) | 11.44 (3) | 14.61 (3) | 15.13 | 15.75 (3) | ||||
| Bio 2 | Annual Mean Diurnal Range¶ (°C) | 3.97 | 11.24 (1,2,3) | 13.50 | 8.19 | ||||
| Bio 3 | Isothermality (%) | 7.04 (1) | 13.20 (2) | 7.70 | 3.35 | ||||
| Bio 4 | Temperature Seasonality (%) | 9.99 | 14.23 | 13.41 (1,2,3) | 7.68 (1,2,3) | ||||
| Bio 5 | Max Temperature of Warmest Month¶ (°C) | 9.08 | 7.58 (1) | 13.31 | 12.28 | ||||
| Bio 6 | Min Temperature of Coldest Month¶ (°C) | 14.87 (2) | 9.82 (2) | 12.36 (1,2,3) | 20.08 (2) | ||||
| Bio 7 | Annual Temperature Range¶ (°C) | 8.18 | 14.33 (1,3) | 14.86 (1,2,3) | 9.24 (1,2,3) | ||||
| Bio 10 | Mean Temperature of Warmest Quarter¶ (°C) | 9.85 (1,2,3) | 9.83 (2) | 14.42 (1,2) | 11.40 | ||||
| Bio 11 | Mean Temperature of Coldest Quarter¶ (°C) | 16.43 (1) | 9.36 (1) | 10.31 (1,2,3) | 18.83 (1) | ||||
| Bio 12 | Annual Precipitation (mm) | 17.59 | 9.01 (1,2,3) | 7.62 (1,2,3) | 13.59 (1,2,3) | ||||
| Bio 13 | Precipitation of Wettest Month (mm) | 14.70 | 7.48 | 4.00 | 5.88 | ||||
| Bio 14 | Precipitation of Driest Month (mm) | 6.00 | 12.70 | 10.54 | 10.43 | ||||
| Bio 15 | Precipitation Seasonality (CV, %) | 9.15 (1,3) | 4.53 (1) | 4.43 (1) | 4.56 | ||||
| Bio 16 | Precipitation of Wettest Quarter (mm) | 15.04 (1,2,3) | 6.56 | 3.54 | 5.99 | ||||
| Bio 17 | Precipitation of Driest Quarter (mm) | 6.68 | 13.88 | 11.10 | 10.80 | ||||
| Criterion/Species | D. mexicanus | P. leiophylla | P. teocote | P. devoniana |
|---|---|---|---|---|
| Calibration and evaluation of candidate models | ||||
| TCM | 1392 | 1392 | 1392 | 1392 |
| SSM | 1383 | 745 | 1328 | 132 |
| MCOr | 262 | 0 | 763 | 201 |
| MAIC | 1 | 3 | 1 | 2 |
| n of SSM and MCOr | 254 | 0 | 763 | 201 |
| n of SSM and MAIC | 1 | 3 | 1 | 2 |
| n of SSM, MCOr and MAIC | 1 | 0 | 1 | 1 |
| Selected model | M_2_F_t (2) | M_5_q (3) | M_3_F_qth (1) | M_2_F_qh (1) |
| Statistics of the selected model | ||||
| Mean AUC ratio | 1.66 | 1.24 | 1.49 | 1.35 |
| Rate of omission > 0.05% | 0.05 | 0.77 | 0.04 | 0.03 |
| AICc | 1683.34 | 17393.42 | 13851.28 | 4900.27 |
| delta AICc | 21.6 | 252.84 | 251.35 | 61.26 |
| Variable | Contrib. | Dendroctonus mexicanus | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | (%) | Length | Mean | MeanCI | 0.05 | 0.10 | 0.25 | Median | 0.75 | 0.90 | 0.95 | Range | SD | CV | MAD | IQR |
| Bio 6 | 4.7 | 86 | 53.0 | ±7.0 | −10.1 | 6.7 | 34.0 | 55.2 | 74.3 | 95.6 | 100.6 | 153.8 | 32.5 | 61.3 | 31.4 | 40.3 |
| Bio 10 | 87.8 | 86 | 180.7 | ±5.6 | 140.5 | 149.6 | 166.3 | 178.7 | 201.3 | 218.5 | 221.2 | 130.7 | 26.1 | 14.4 | 28.1 | 35.0 |
| Bio 15 | 3.2 | 86 | 89.2 | ±2.7 | 65.9 | 71.8 | 79.8 | 90.7 | 100.2 | 103.4 | 106.4 | 52.6 | 12.6 | 14.1 | 14.3 | 20.4 |
| Bio 16 | 4.3 | 86 | 578.2 | ±43.7 | 307.8 | 325.0 | 430.5 | 546.0 | 708.8 | 877.0 | 962.0 | 834.0 | 204.0 | 35.3 | 201.6 | 278.3 |
| Pinus leiophylla | ||||||||||||||||
| Bio 1 | 93.9 | 900 | 138.7 | ±1.2 | 111.9 | 116.1 | 126.5 | 136.4 | 149.5 | 164.1 | 171.4 | 99.0 | 17.9 | 12.9 | 17.1 | 23.0 |
| Bio 2 | 5.6 | 900 | 118.9 | ±0.7 | 95.1 | 103.1 | 116.8 | 123.0 | 125.7 | 128.1 | 129.0 | 62.3 | 11.0 | 9.3 | 4.9 | 9.0 |
| Bio 7 | 0.5 | 900 | 246.6 | ±2.8 | 169.3 | 181.2 | 219.9 | 253.9 | 277.4 | 297.1 | 305.8 | 202.2 | 42.1 | 17.1 | 40.5 | 57.5 |
| Pinus teocote | ||||||||||||||||
| Bio 4 | 3.8 | 735 | 3363.6 | ±79.6 | 1195.5 | 1553.3 | 2962.3 | 3628.8 | 3956.4 | 4698.7 | 4916.2 | 533.6 | 1098.5 | 32.7 | 600.7 | 994.1 |
| Bio 6 | 15.6 | 735 | 18.4 | ±2.7 | −19.5 | −14.6 | −6.4 | 3.5 | 37.0 | 78.1 | 93.9 | 195.5 | 36.6 | 198.9 | 20.1 | 43.4 |
| Bio 7 | 3.3 | 735 | 227.9 | ±2.8 | 156.2 | 164.3 | 212.2 | 238.3 | 251.5 | 269.1 | 275.3 | 200.0 | 38.0 | 16.7 | 23.1 | 39.3 |
| Bio 10 | 63.8 | 735 | 178.5 | ±1.3 | 152.6 | 158.2 | 166.7 | 176.0 | 187.9 | 201.9 | 210.9 | 150.3 | 18.5 | 10.4 | 14.9 | 21.1 |
| Bio 11 | 5 | 735 | 87.6 | ±2.3 | 51.8 | 57.3 | 66.1 | 76.9 | 102.8 | 138.5 | 153.6 | 183.4 | 31.8 | 36.3 | 20.6 | 36.6 |
| Bio 12 | 6.5 | 735 | 901.0 | ±18.0 | 581.7 | 626.0 | 723.0 | 847.0 | 1045.5 | 1250.6 | 1392.9 | 1424.0 | 248.5 | 27.6 | 225.4 | 322.5 |
| Bio 15 | 2.1 | 735 | 92.1 | ±0.9 | 71.4 | 76.5 | 83.9 | 92.9 | 101.1 | 106.5 | 110.3 | 76.0 | 11.8 | 12.8 | 12.9 | 17.2 |
| Pinus devoniana | ||||||||||||||||
| Bio 4 | 28.2 | 255 | 1813.2 | ±61.3 | 1072.3 | 1165.8 | 1523.7 | 1780.2 | 1985.4 | 2440.0 | 2745.9 | 2892.5 | 497.3 | 27.4 | 338.6 | 461.7 |
| Bio 7 | 1.3 | 255 | 181.9 | ±3.2 | 132.0 | 139.0 | 162.7 | 189.6 | 199.1 | 212.2 | 218.3 | 115.3 | 26.1 | 14.3 | 23.4 | 36.4 |
| Bio 11 | 50.8 | 255 | 141.9 | ±3.5 | 99.8 | 107.2 | 119.3 | 139.7 | 162.0 | 176.8 | 188.3 | 140.5 | 27.5 | 19.4 | 31.7 | 42.7 |
| Bio 12 | 19.8 | 255 | 1090.0 | ±40.5 | 608.8 | 712.4 | 858.5 | 1043.0 | 1265.5 | 1579.4 | 1770.8 | 1624.0 | 328.7 | 30.2 | 309.9 | 407.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Encina, F.M.; Méndez-González, J.; Mendieta-Oviedo, R.; López-Díaz, J.Ó.M.; Nájera-Luna, J.A. Ecological Niches and Suitability Areas of Three Host Pine Species of Bark Beetle Dendroctonus mexicanus Hopkins. Forests 2021, 12, 385. https://doi.org/10.3390/f12040385
Méndez-Encina FM, Méndez-González J, Mendieta-Oviedo R, López-Díaz JÓM, Nájera-Luna JA. Ecological Niches and Suitability Areas of Three Host Pine Species of Bark Beetle Dendroctonus mexicanus Hopkins. Forests. 2021; 12(4):385. https://doi.org/10.3390/f12040385
Chicago/Turabian StyleMéndez-Encina, Fátima M., Jorge Méndez-González, Rocío Mendieta-Oviedo, José Ó. M. López-Díaz, and Juan A. Nájera-Luna. 2021. "Ecological Niches and Suitability Areas of Three Host Pine Species of Bark Beetle Dendroctonus mexicanus Hopkins" Forests 12, no. 4: 385. https://doi.org/10.3390/f12040385