
Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland


Plants and Pathogens Group, Institute Land Nature Environment, Geneva School of Engineering Architecture and Landscape (HEPIA), HES-SO University of Applied Sciences and Arts Western Switzerland, 150 Route de Presinge, 1254 Jussy, Switzerland
*
Author to whom correspondence should be addressed.
Forests 2021, 12(4), 434; https://doi.org/10.3390/f12040434
Submission received: 15 February 2021
/
Revised: 22 March 2021
/
Accepted: 31 March 2021
/
Published: 4 April 2021
(This article belongs to the Special Issue Pests and Pathogens of Urban Trees)
Abstract
:Fungi were isolated in pure cultures from decaying giant sequoias in Geneva (Switzerland). Isolates were genetically identified by ITS rDNA sequencing. Young giant sequoia trees were artificially infected with a pure culture of Botryosphaeria parva. Henle-Koch’s Postulates demonstrated that Botryosphaeria parva was pathogenic to Sequoiadendron giganteum. When analysing the microorganisms associated to canker and dieback symptoms in a giant sequoias (Sequoiadendron giganteum) in Geneva, the fungus Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips, teleomorph Botryosphaeria parva (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips, was isolated, whereas such symptoms are commonly associated to Fusicoccum aesculi (teleomorph Botryosphaeria dothidea). These two fungal species belong to the same genus Botryosphaeria of the Botryosphaeriaceae family. Because Neofusicoccum parvum was causing cankers and diebacks in other woody species around the world, we extended the analysis to other trees displaying sequoia dieback symptoms in order to evaluate the involvement of Neofusicoccum parvum in such increasing symptoms in sequoias in Geneva. Dried twigs, trunk, and branch cankers from diseased trees were sampled on several distinct sites. From all samples, isolated fungi in pure cultures showed a phenotype typical of Botryosphaeriaceae species. Isolates were then genetically identified at the species level. Subsequently Neofusicoccum parvum was inoculated to young giant sequoia trees, re-isolated in pure culture from provoked symptoms, and re-identified to fulfil Henle-Koch’s postulates. The identification confirmed that Neofusicoccum parvum was present on all sites, while Fusicoccum aesculi was retrieved only once alone. The inoculation of Neofusicoccum parvum isolates on young sequoias demonstrated for the first time that this fungus was able to develop cankers in Sequoiadendron giganteum. Neofusicoccum parvum could, therefore, be the major cause for dying of giant sequoias in the Geneva Lake area.
1. Introduction
Introduced in the 1850s to Europe, giant sequoia trees (Sequoiadendron giganteum (Lindl.) J. Buchholz, 1939) met a general and genuine craze. The first sequoia trees were also sewed and planted at the same time in Geneva. Some of the still-living trees today date from this period. Most of the 350 giant sequoias, present in the territory of the canton of Geneva, now show symptoms of a disease similar to that caused by the fungal pathogen Fusicoccum aesculi [1]. The fungus is also known under the name of its teleomorph Botryosphaeria dothidea (Moug. ex Fr.) Ces. & De Not., [2] (Botryosphaeriaceae). This fungus is known to cause the Botryosphaeria canker and white rot on giant sequoia trees of Sequoiadendron giganteum, but also on Sequoia sempervirens and incense cedars Calocedrus decurrens in their native region [3,4,5]. Its pathogenicity was described on S. giganteum and S. sempervirens [4]. Symptoms are known to be more acute on trees established in non-native regions than in the regions of origin. This is especially true in hot and dry climates, where the disease usually leads to trees diebacks [6,7]. Botryosphaeria dothidea has been known for 150 years as a mild pathogenic fungus and a common cause of canker and dieback of trees worldwide on 24 host genera [8], on which it may provoke cankers on twigs, branches, and stems, resulting in tips and branches dieback and finally plant death. Other symptoms on some tree species may include fruit rot and blue stains [8]. It is now also known to be usually present as a latent endophytic pathogen, which expresses its pathogenicity in plant stress conditions [8]. Recently, another fungus of the same Botryosphaeriaceae family, named Neofusicoccum parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips [9,10] (teleomorph: Botryosphaeria parva (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips) [10] was found in studies of Botryosphaeria canker on giant sequoias in Greece [11]. These two cosmopolitan fungal species are, however, widespread, in a very large range of plant hosts, as endophytes as well as pathogens [12,13,14]. If B. dothidea has long been considered a marginally pathogenic fungus, N. parvum seems to be more aggressive on important crops such as grapevine [15] and olive trees [16]. On giant sequoias, the symptoms associated to an infection by these Botryosphaeria species include the shrivelling of twigs of the crown, distributed in patches, and the appearance of cankers on twigs, branches, and trunks, associated with resin flows. Early trunk cankers are discrete in reason of the thickness of the bark of sequoias. With the progress of the infection, the crown displays a discoloured foliage and is naturally pruned from dead twigs and branches. In this study, we have isolated Botryosphaeriaceae strains from diseased giant sequoia trees in different locations of the Canton of Geneva, in order to isolate and accurately identify the pathogen, which has been causing these dieback and canker symptoms resulting in an increased mortality of these trees in recent years.
2. Materials and Methods
2.1. Sampled Trees
2.2. Isolation of Microorganisms
Small twigs were collected in the proximity of a canker on a branch and cut in pieces 3–4 cm long. After successive surface disinfection and washing steps, according to Slippers and Wingfield [12], these twig pieces were left to air dry under a laminar flow. More precisely, disinfection consisted in 30 s in ethanol 70% (v/v), followed by 1 min in NaOCl 1% (w/v), 30 s in ethanol 70% (v/v), and was terminated by washing with sterile ultrapure water. Twig fragments were then sampled at the border of the infected area and put on the surface of Potato Glucose Agar (PGA) medium (Carl Roth, Switzerland) supplemented with Ampicillin at 100 mg/L final concentration in Petri dishes, and then incubated at room temperature. From two individual trees, supplementary samples were also collected from black fungal stroma observed on the trunk bark and directly inoculated on PGA Ampicillin medium and incubated in the same conditions.
2.3. Genetic Identification
All colonies displaying a culture pattern typical of Botryosphaeriaceae species (Figure 4a–c) were isolated in pure cultures and sub-cultured in 50 mL Potato Glucose Broth (PGB) (Carl Roth, Switzerland) liquid medium for 3–4 days. Resulting mycelium was ground in liquid nitrogen and DNA was extracted following a protocol adapted from Lefort and Douglas [17]. DNA concentrations were measured in a spectrometer Nanodrop ND-100 (Wilmington, DE, USA). Finally, all samples had their DNA concentration adjusted to 50 ng/µL in sterile ultrapure water. The internal transcribed spacer (ITS) region was amplified using the primer pair ITS4/ITS5 [18] synthesised at Microsynth AG (Balgach, Switzerland). Polymerase Chain Reaction (PCR) amplifications were carried out in a T Professional Basic Gradient thermocycler (Biometra, Göttingen, Germany). PCR reaction mixtures contained 15 ng template DNA in 1× PCR buffer (Biotools, Madrid, Spain), 2 mM MgCl2, 0.5 µM of each primer, 100 µM of dNTPs (Sigma Aldrich, Switzerland), and 0.5 units of Taq DNA polymerase (Biotools, Spain) in a reaction volume of 25 µL. The applied PCR program was as follows: an initial denaturation of 3 min at 95 °C, followed by 38 cycles of 60 s at 95 °C, 45 s at 56 °C, and 60 s at 72°C, terminated by a final elongation for 6 min at 72 °C. PCR products were analysed on 1% agar gel in 1% TBE buffer (Tris-HCL 89 mM, boric acid 89 mM, EDTA 2 mM, pH 8), purified with a Wizard® PCR Clean-Up System (Promega, Dübendorf, Switzerland) and assayed as described previously. Sanger DNA sequencing was performed at Microsynth AG.
2.4. Production of Pycnidia and Measurement of Conidia Dimensions
In order to obtain fungal fructifications, pure cultures from all retained strains were inoculated on H2O-agar 2% Petri plates. Pieces of grapevine petioles were surface sterilised following a published protocol [19], and then deposited on these inoculated Petri plates. Cultures were then incubated for 3 to 4 weeks at 25 ± 2 °C in a Percival (Perry, IA, USA) climatic chamber under fluorescent light and under a day-night regime (12:12). When pycnidia were fully developed, they were dissected and observed under a binocular stereoscopic microscope Motic SMZ168 Series (Motic Inc. LTD, Kowloon Bay, Hong-Kong) fitted with a camera Axiomcam ERc5s (Zeiss, Jena, Germany), in order to obtain free conidia. Conidia measurements were carried out under a dissecting microscope Axiostar HBO50/AC (Zeiss) using the software Axiovision LE on pictures obtained with the camera Axiomcam MRc (Zeiss).
2.5. Pathogenicity Testing According Henle-Koch’s Postulates
In order to ascertain the hypothesis according to which the isolated organism was responsible for the observed symptoms on giant sequoias, the experiments of the Henle-Koch’s postulates were implemented [20,21]. Young and healthy individuals of Sequoiadendron giganteum (60 cm high) were kindly provided potted by the ornamental orchards of the Centre for Professional Training (CFPNE) of Lullier. Trees and pots were washed off dust with water and then briefly surface disinfected with 70% ethanol. Trees were then cultivated in a climatic chamber for four months, with appropriate watering, under the following conditions: 16 h light/8 h dark photoperiod with a light intensity of 6000 lux provided by cool white fluorescent (Sylvania LUXLINE PLUS F58W/840-T8, 1500 mm) and purple photosynthetic lamps (Sylvania GROLUX F58W/GRO, 1500 mm). Two trees were kept as the negative control and two trees were inoculated with a fresh fungal culture on PGA (5 d) of Neofusicoccum parvum. Inoculation was performed by punching a piece of bark (4 mm diam., 2 mm depth) and filling the wound with sterile PGA for controls or Neofusicoccum parvum culture for the test trees, with the help of a sterile scalpel. In all trees, the wounds were protected by ParafilmTM to avoid desiccation. As three trees (1 control and 2 test trees), out of four, were forked trees, a second inoculation was performed on the second branch. This yielded four pathogen inoculations on two test trees and three water inoculations on two control trees in total. The experiment was led until cankers developed. Molecular identification by gene amplification and sequencing, as described previously, was used to identify the pathogen after re-isolating from cankers for comparison with the inoculated pathogen.
3. Results
3.1. Isolation and Genetic Identification
Sequences obtained for PCR products were registered in GenBank under the Accessions number MH500822 to MH500841. These sequences were compared with the sequences of the GenBank database of the National Centre for Biotechnology Information [22] using the BLASTn Tool (NCBI) [23]. These sequences shared 99–100% identity with the ITS sequence of the type strain ICM 8003/CMV 9081 (GenBank Accession NR_119487) [1] over 99% sequence length and with the 40 first matching sequences of Neofusicoccum parvum strains (99.62–100% identities, over 99–100% of their sequences). On the other hand, they only shared around 93% identity in average over 96% of the ITS sequence of the type strain CMV 8000 of Fusicoccum aesculi [1]. As shown on Table 1, Neofusicoccum parvum was isolated in eight trees out nine sampled trees, in seven distant locations, whereas Fusicoccum aesculi (Botryosphaeria dothidea) was only retrieved alone from one sequoia tree, slightly affected, in one location, and from another tree in combination with Neofusicoccum parvum in another location. Only one tree (S. giganteum “Glauca” on the site of Lullier) did not reveal any Botryosphaeriaceae, but a Sordaria sp. isolate instead, although it displayed a few dry twigs and little cankers on branches. Another sequoia tree located in Lullier, S. giganteum “Pendulum,” showed Neofusicoccum parvum on diseased twigs presenting canker symptoms, another Botryosphaeriaceae species, Diplodia mutila, anamorph of Botryosphaeria stevensii, on its bark. This last species is known to colonise barks of ash trees [24], grapevine [25], and London plane trees [26]. It is noticeable that Neofusicoccum parvum has been isolated from the seven studied sites. Isolates of Neofusicoccum parvum, Fusicoccum aesculi (Botryosphaeria dothidea), and Diplodia mutila (Botryosphaeria stevensii) are shown in Figure 4a–c.
3.2. Pycnidia Production and Conidia Dimensions
Growing the two possible agents of redwood canker, Neofusicoccum parvum and Fusicoccum aesculi, on the surface of grapevine twigs yielded similar pycnidia for the two species as shown on Figure 4d (Pycnidia of Neofusicoccum parvum) and Figure 4e (Pycnidia of Fusicoccum aesculi). Conidia of Neofusicoccum parvum were single cells, hyaline, spindle-shaped to ellipsoidal, with thin cell walls (see Figure 4f–g) They displayed a rounded apex with a truncated or rounded basis. The conidia measured 12.1–16.9 µm (average 14.3 ± 1.3 µm) in length and 3.9–5.5 µm (average 4.7 ± 0.4 µm) in width, which is congruent with previous reports [27].
3.3. Pathogenicity Test
The testing system constituted of two control trees and two inoculated trees is shown on Figure 5a,b. After only one week, necrosis was visible on twigs inoculated with Neofusicoccum parvum. In less than 20 d, the branch above and below the inoculation point was dried and black (Figure 5d,e), whereas the same branch of the negative control remained very green and healthy (Figure 5c). Necrosis appeared later on the trunks of inoculated trees. After 30 days, while control trees were closing the wounds with a cicatricial bulge (Figure 6a–b), inoculated trees hardly succeeded in creating this cicatricial bulge and bark scales were already desiccated. After 4 months, cankers were visible in place of wounds on inoculated trees: bark discoloration, desquamating bark scales, and trunk girdling (Figure 6c–d). Pure cultures re-isolated from cankers of the two inoculated trees were then all genetically identified as belonging to the species Neofusicoccum parvum.
4. Discussion
In contrary to what has been thought by inference for the last 30 years, the main pathogenic agent of dieback of giant sequoias, at least in Geneva, would not only be Fusicoccum aesculi (Botryosphaeria dothidea), but a close fungus of the species Neofusicoccum parvum (Botryosphaeria parva) could also be responsible for the same symptoms and, therefore, for the same disease. This fungus, when obtained in pure culture, quickly reproduces dieback symptoms in climatic chambers after inoculation to young healthy giant sequoia individuals. These results confirm previous observations in Greece [9], where this species was reported as a causing agent of giant sequoias dieback, though without Henle- Koch’s postulates implementation. We report here the first evidence of Neofusicoccum parvum (Botryosphaeria parva) being pathogenic to Sequoiadendron giganteum trees, as shown unambiguously by the results of the Henle-Koch’s Postulates. This observation of Neofusicoccum parvum being a woody plant pathogen is congruent with the report of similar symptoms observed on Platanus × acerifolia trees in Geneva. These symptoms were attributed to this fungal species after demonstrating that this fungal species was causing cankers in London plane trees [26].
Several hypotheses can be formulated concerning the pathogenicity of this fungus. First, Neofusicoccum parvum (Botryosphaeria parva) was always, in fact, the causing agent of the symptoms observed on giant sequoias in Geneva. This disease recently appeared in Geneva, and was for a long time likely mistaken with Fusicoccum aesculi (Botryosphaeria dothidea) because it is morphologically very close under microscopical observation from this fungus. Additionally, the symptoms are similar. Pycnidia and conidia from both species are also very similar as shown in the present study. Furthermore, Fusicoccum aesculi (Botryosphaeria dothidea) was first reported as Botryosphaeria ribis, the pathogenic agent on sequoias [3], branded as the causing agent of giant sequoias cankers, without performing Henle-Koch’s Postulates. This disease seems to have already been common outside the native region of giant sequoias in this time [3,4,5]. It was shown a short while later that Botryosphaeria dothidea (or B. ribis) was pathogenic to Sequoiadendron giganteum and Sequoia sempervirens [4]. In this case, Henle-Koch’s Postulates were performed, starting from an inoculum from a twig, which was morphologically described as Botryosphaeria dothidea, and thought to be, at this time, a synonym of Botryosphaeria ribis. B. ribis [28], now known as Neofusicoccum ribis [29], is genetically closed to N. parvum [29], while B. dothidea is genetically very distinct [29,30]. Based on the initial work and on the symptoms described [4], cankers and diebacks of Sequoiadendron giganteum are generally attributed to Botryosphaeria dothidea without further expertise by genetic identification. Besides the fact that it is genetically very distinct from N. parvum [29] and N. ribis [30], its conidia are much bigger [30]. B. dothidea is also very polymorphic and has been described as containing two to three genetic groups based on ITS sequences, with diverse pathogenic properties in relation to specific hosts [31,32,33,34].
More precise techniques of genetic identification now allow us to identify without any ambiguity Neofusicoccum parvum (Botryosphaeria parva), which is sufficiently different of Fusicoccum aesculi (Botryosphaeria dothidea), in the genetic sequence of the taxonomic region of the ribosomal internal transcribed spacer (ITS). It is close of Neofusicoccum ribis, at such a point that many authors report a N. ribis/N. parvum species complex [35,36], but is still genetically distinguishable. Such a level of discrimination, nowadays routinely achieved, leads to a better description of several pathogenic Botryosphaeriaceae species affecting giant sequoias. For instance, two species were reported in the USA, each on one occasion, on giant sequoias; these are Neofusicoccum mediterraneum [37] and Neofusicoccum nonquaesitum [38]. A third one has been reported in Australia and named Neofusicoccum australis [39]. These three species have not been observed in Switzerland yet. Another alternative hypothesis could be that Neofusicoccum parvum (Botryosphaeria parva) is a novel-causing agent of dieback in giant sequoias, of which a more aggressive strain, more pathogenic, and more adapted to new climatic conditions, and therefore more competitive, could have replaced the other species Fusicoccum aesculi or B ribis. Climatic conditions also appear to affect giant sequoia trees’ health as hotter droughts were reportedly linked to foliage dieback [40]. Recently, the genetic basis of local adaptation of giant sequoias to summer precipitations was unveiled and revealed variability in genomic regions correlated to precipitations and temperature [41]. This would explain the observed behaviour and sensibility to pathogens of giant sequoias in climate change. Identifying genes linked to local adaptation of giant sequoia to climate change could, therefore, represent a selection tool for better-adapted giant sequoias to local conditions. Finally, a third hypothesis would be that Neofusicoccum parvum (Botryosphaeria parva) could also be an endophytic fungus of giant sequoias and that imbalances in endophytic microbiota, under changing climatic conditions, would repress or remove another member of this microbiota, which was controlling Neofusicoccum parvum, allowing it to develop its pathogenicity. This last hypothesis was not supported by our trials of cultivation and isolation from the cultivable microbiota from a giant sequoia branch, in which we did not find any Botryosphaeriaceae species (data not shown), but such preliminary works should be extended.
5. Conclusions
Neofusicoccum parvum (Botryosphaeria parva) was found as predominantly present on diseased samples from giant sequoias in Geneva, while Fusicoccum aesculi (Botryosphaeria dothidea) was retrieved only once alone. The inoculation of Neofusicoccum parvum isolates on young sequoias followed by its re-isolation and genetic identification from diseased tissues showed for the first time that this fungus was able to develop cankers in Sequoiadendron giganteum. This leads us to think that B. dothidea was possibly initially misidentified on giant sequoias and that Neofusicoccum parvum could, therefore, be the major cause for giant sequoias cankers and dieback.
Author Contributions
Conceptualization, M.H. and F.L.; Data curation, F.L.; Formal analysis, M.H., B.C. and J.C.; Funding acquisition, F.L.; Investigation, M.H., B.C., J.C., R.C. and F.L.; Methodology, M.H., J.C. and F.L.; Project administration, F.L.; Resources, R.C.; Supervision, F.L.; Validation, F.L.; Writing—original draft, M.H. and F.L.; Writing—review & editing, M.H. and F.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the State directorate for Agriculture and Nature (OCAN) of the Canton and Republic of Geneva and the strategic research fund of the HES-SO University of Applied Sciences and Arts Western Switzerland.
Acknowledgments
We are particularly grateful Roger Beer and Bertrand Favre, from OCAN, for their continuous support and to Gilles Villanova, responsible of the ornamental orchards of the Centre for Professional Training Nature Environment (CFPNE) of Lullier, for providing young sequoia trees.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Slippers, B.; Crous, P.W.; Denman, S.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Combined multiple gene genealogies and phenotypic characters differentiate several species previously identified as Botryosphaeria dothidea. Mycologia 2004, 96, 83–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesati, V.; De Notaris, G. Schema di classificazione degle sferiacei italici aschigeri piu’ o meno appartenenti al genere Sphaeria nell’antico significato attribuitoglide Persono. Comm. Soc. Critt. Ital. 1863, 1, 177–420. [Google Scholar]
- Bega, R.V. Diseases of Pacific Coast Conifers. Agriculture Handbook No 521; Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1978; p. 75. [Google Scholar]
- Worrall, J.J.; Correll, J.C.; McCain, A.H. Pathogenicity and teleomorph-anamorph connection of Botryosphaeria dothidea on Sequoiadendron giganteum and Sequoia sempervirens. Plant Dis. 1986, 70, 757–759. [Google Scholar] [CrossRef]
- Wood, D.L. Pests of the Native California Conifers. In California Natural History Guides; University of California Press: Oakland, CA, USA, 2003. [Google Scholar]
- Kehr, R. Triebschäden an Mammutbaum (Sequoiadendron giganteum) durch Botryosphaeria dothidea auch in Deutschland nachgewiesen. Nachrichtenblatt des Deutschen Pflanzenschutzdienstes 2004, 56, 37–43. [Google Scholar]
- Cech, T.; Tomiczek, C. Botryosphaeria-Krebs (Botryosphaeria dothidea) des Mammutbaumes nimmt in Ostösterreich zu. Forstsch. Aktuell 2013, 57/58, 37–40. [Google Scholar]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea, a latent pathogen of global importance to woody plant health. Mol. Plant Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef]
- Pennycook, S.R.; Samuels, G.J. Botryosphaeria and Fusicoccum species associated with ripe fruit rot of Actinidia deliciosa (kiwifruit) in New Zealand. Mycotaxon 1985, 24, 445–458. [Google Scholar]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Philips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Tsopelas, B.; Slippers, B.; Wingfield, M.J.; Gonou-Zagou, Z. Infection of Sequoiadendron giganteum in Greece by the fungus Neofusicoccum parvum. Phytopathol. Mediterr. 2009, 48, 311. [Google Scholar]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants, diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–108. [Google Scholar] [CrossRef]
- Golzar, H.; Burgess, T.I. Neofusicoccum parvum, a causal agent associated with cankers and decline of Norfolk Island pine in Australia. Austral. Plant Pathol. 2011, 40, 484–489. [Google Scholar] [CrossRef]
- Park, S.; Kim, S.-H.; Back, C.-G.; Lee, S.-Y.; Kang, I.-K.; Jung, H.-Y. First Report of Botryosphaeria parva causing stem blight on Rubus crataegifolius in Korea. Res. Plant Dis. 2016, 22, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Massonnet, M.; Figueroa-Balderas, R.; Galarneau, E.R.A.; Miki, S.; Lawrence, D.P.; Sun, Q.; Wallis, C.M.; Baumgartner, K.; Cantu, D. Neofusicoccum parvum colonization of the grapevine woody stem triggers asynchronous host responses at the site of infection and in the leaves. Front. Plant Sci. 2017, 8, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.; Raimondo, M.; Cibelli, F.; Phillips, A.; Lops, F. Pleurostomophora richardsiae, Neofusicoccum parvum and Phaeoacremonium aleophilum; associated with a decline of olives in Southern Italy. Phytopathol. Mediterr. 2013, 52, 517–527. [Google Scholar] [CrossRef]
- Lefort, F.; Douglas, G.C. An efficient micro-method of DNA isolation from mature leaves of four hardwood tree species Acer, Fraxinus, Prunus and Quercus. Ann. For. Sci. 1999, 56, 259–263. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Van Niekerk, J.M.; Crous, P.W.; Groenewald, J.Z.; Fourie, P.H.; Halleen, F. DNA phylogeny, morphology and pathogenicity of Botryosphaeria species on grapevines. Mycologia 2004, 96, 781–798. [Google Scholar] [CrossRef]
- Henle, J. Pathologische Untersuchungen—Von den Miasmen und Contagien und von den Misamatisch-Contagiösen Krankheiten; Verlag von August Hirschwald: Berlin, Germany, 1840. [Google Scholar]
- Koch, R. Die Aetiologie der Tuberculose. Mitt. Kaiser. Gesundh. 1884, 2, 1–88. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [Green Version]
- Meier, F.; Engesser, R.; Forster, B.; Odermatt, O.; Angst, A. Protection des Forêts—Vue D’ensemble 2009; Institut fédéral de Recherches sur la Forêt, la Neige et le Paysage WSL: Birmensdorf, Switzerland, 2010. [Google Scholar]
- Lehoczky, J. Black dead-arm disease of grapevine caused by Botryosphaeria stevensii infection. Acta Phytopathol. Acad. Sci. Hung. 1974, 9, 319–327. [Google Scholar]
- Pelleteret, P.; Crovadore, J.; Cochard, B.; Pasche, S.; Bovigny, P.-Y.; Chablais, R.; Lefort, F. Urban London plane tree dieback linked to fungi in the Botryosphaeriaceae. Urban Urban Green 2017, 22, 74–83. [Google Scholar] [CrossRef]
- Haleem, R.A.; Abdullah, S.K.; Jubrael, J.M.S. Identification and pathogenicity of Botryosphaeria parva associated with grapevine decline in Kurdistan region—Iraq. Acta Agrobot. 2012, 65, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Grossenbacher, J.G.; Duggar, B.M. A contribution to the life history, parasitism, and biology of Botryosphaeria ribis. N. Y. Agric. Exp. Stn. 1911, 18, 114–188. [Google Scholar]
- Phillips, A.J.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.; Stanosz, G. Molecular and morphological differentiation of Botryosphaeria dothidea (anamorph Fusicoccum aesculi) from some other fungi with Fusicoccum anamorphs. Mycologia 2001, 93, 505–515. [Google Scholar] [CrossRef]
- Smith, D.R.; Michailides, T.J.; Stanosz, G.R. Differentiation of a Fusicoccum species causing panicle and shoot blight on California pistachio trees from Botryosphaeria dothidea. Plant Dis. 2001, 85, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Michailides, T.J. Characterization of Botryosphaeria dothidea isolates collected from pistachio and other plant hosts in California. Phytopathology 2002, 92, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Denman, S.; Crous, P.W.; Taylor, J.E.; Kang, J.; Pascoe, I.; Wingfield, M.J. An overview of the taxonomic history of Botryosphaeria and a re-evaluation of its anamorphs based on morphology and ITS rDNA phylogeny. Stud. Mycol. 2000, 45, 129–140. [Google Scholar]
- Ogata, T.; Sano, T.; Harada, Y. Botryosphaeria spp. isolated from apple and several deciduous fruit trees are divided into three groups based on the production of warts on twigs, size of conidia, and nucleotide sequences of nuclear ribosomal DNA ITS regions. Mycoscience 2000, 41, 331–337. [Google Scholar] [CrossRef]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Multiple gene genealogies and phenotypic data reveal cryptic species of the Botryosphaeriaceae: A case study on the Neofusicoccum parvum/N. ribis complex. Mol. Phylogenet. Evol. 2009, 51, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Pavlic, D.; Slippers, B.; Coutinho, T.A.; Wingfield, M.J. Molecular and phenotypic characterisation of three phylogenetic species discovered within the Neofusicoccum parvum/N. ribis complex. Mycologia 2009, 101, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Inderbitzin, P.; Bostock, R.M.; Trouillas, F.P.; Michailides, T.J. A six-locus phylogeny reveals high species diversity in Botryosphaeriaceae from California almond. Mycologia 2010, 102, 1350–1368. [Google Scholar] [CrossRef]
- Rooney-Latham, S.; Tidwell, T.E.; Blomquist, C.L.; Peek, K.S. First report of Neofusicoccum nonquaesitum causing branch cankers on giant sequoia (Sequoiadendron giganteum) in North America. Plant Dis. 2012, 96, 905. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Multiple gene sequences delimit Botryosphaeria australis sp. nov. from B. lutea. Mycologia 2004, 96, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, N.L.; Das, A.J.; Ampersee, N.J.; Cahill, K.G.; Caprio, A.C.; Sanders, J.E.; Williams, A.P. Patterns and correlates of giant sequoia foliage dieback during California’s 2012–2016 hotter drought. For. Ecol. Manag. 2018, 419–420, 268–278. [Google Scholar] [CrossRef]
- DeSilva, R.; Dodd, R.S. Association of genetic and climatic variability in giant sequoia, Sequoiadendron giganteum, reveals signatures of local adaptation along moisture-related gradients. Ecol. Evol. 2020, 10, 10619–10632. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A giant sequoia displaying symptoms caused by a Botryosphaeria sp.

Figure 2.
Symptoms of dieback and cankers caused by a Botryosphaeria sp. on diverse giant sequoia trees in Geneva. (a) Shrivelling of a twig; (b) Scattered patches of diseased twigs in the crown; (c) Cross section in a canker showing internal wood necrosis; (d) Branch cankers; (e,f) Resin dripping out of infected twigs.
Figure 2.
Symptoms of dieback and cankers caused by a Botryosphaeria sp. on diverse giant sequoia trees in Geneva. (a) Shrivelling of a twig; (b) Scattered patches of diseased twigs in the crown; (c) Cross section in a canker showing internal wood necrosis; (d) Branch cankers; (e,f) Resin dripping out of infected twigs.

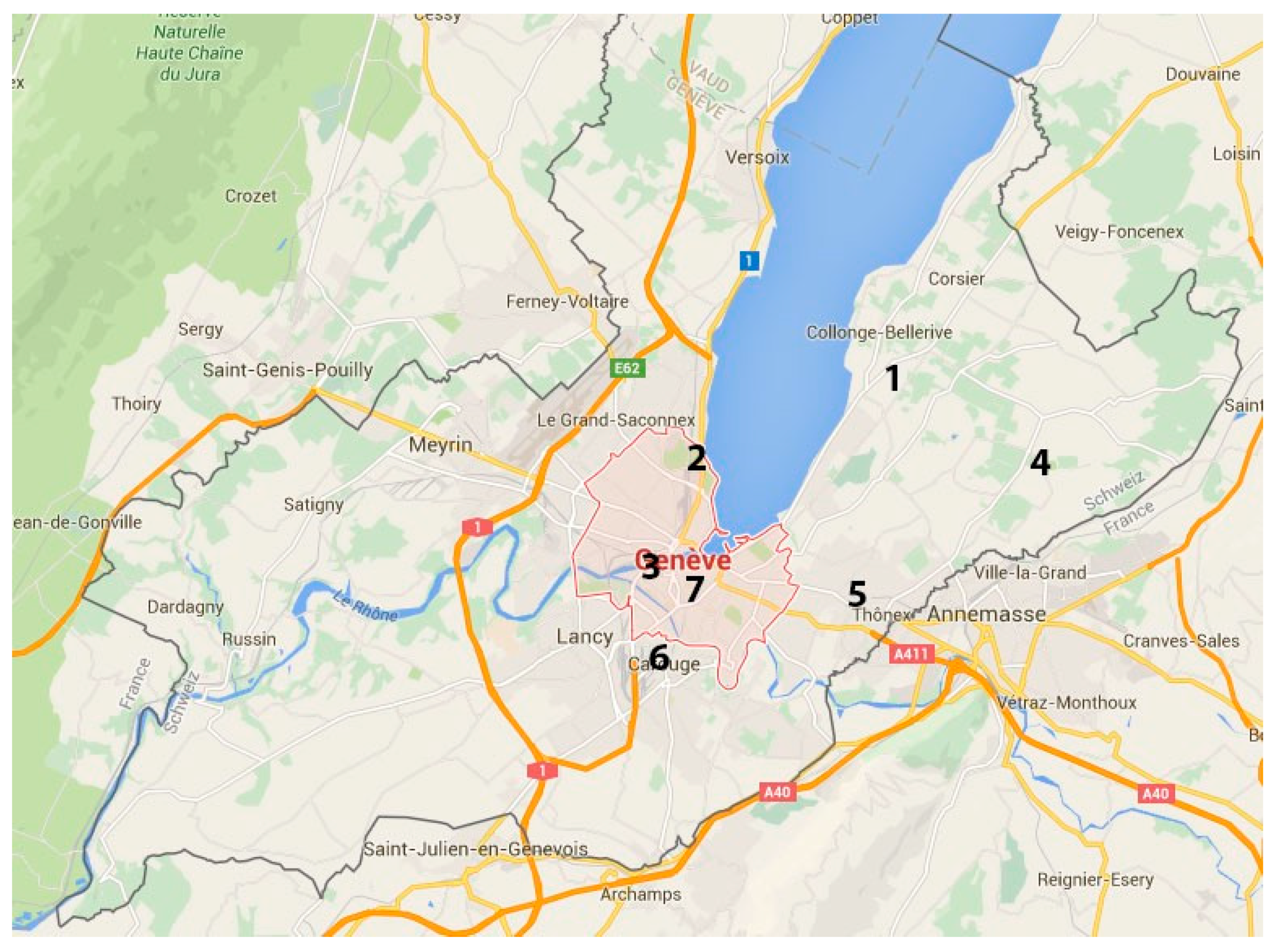
Figure 3.
Localisation of sampled trees: 1. Vésenaz. 2. Geneva (Parc Barton). 3. Geneva (Cimetière des Rois). 4. Lullier. 5. Chêne-Bourg (Parc de l’Ancienne Mairie). 6. Carouge (Boulevard des Promenades). 7. Geneva (Parc des Bastions).
Figure 3.
Localisation of sampled trees: 1. Vésenaz. 2. Geneva (Parc Barton). 3. Geneva (Cimetière des Rois). 4. Lullier. 5. Chêne-Bourg (Parc de l’Ancienne Mairie). 6. Carouge (Boulevard des Promenades). 7. Geneva (Parc des Bastions).

Figure 4.
(a–c): Appearance of pure Botryosphaeriaceae colonies on PGA ampicillin after 10 d incubation at room temperature. (a) Neofusicoccum parvum (Botryosphaeria parva); (b) Fusicoccum aesculi (Botryosphaeria dothidea); (c) Diplodia mutila (Botryosphaeria stevensii); (d,e) Pycnids at the surface of grapevine twigs; (d) Neofusicoccum parvum; (e) Fusicoccum aesculi; (f,g) Conidia of Neofusicoccum parvum (×400) after staining with lactophenol blue. Scale bars (f,g) = 10 µm.
Figure 4.
(a–c): Appearance of pure Botryosphaeriaceae colonies on PGA ampicillin after 10 d incubation at room temperature. (a) Neofusicoccum parvum (Botryosphaeria parva); (b) Fusicoccum aesculi (Botryosphaeria dothidea); (c) Diplodia mutila (Botryosphaeria stevensii); (d,e) Pycnids at the surface of grapevine twigs; (d) Neofusicoccum parvum; (e) Fusicoccum aesculi; (f,g) Conidia of Neofusicoccum parvum (×400) after staining with lactophenol blue. Scale bars (f,g) = 10 µm.

Figure 5.
Pathogenicity testing according to Henle-Koch’s Postulates. (a) Young sequoias after inoculation; (b) Example of an inoculated branch.; (c) Control with inoculation wound (arrow) 20 d after inoculation. (d,e) Young sequoias inoculated with a pure culture of Neofusicoccum parvum 20 d after inoculation, with dieback of the whole twig above and below the inoculation point. Arrows indicate the inoculation point.
Figure 5.
Pathogenicity testing according to Henle-Koch’s Postulates. (a) Young sequoias after inoculation; (b) Example of an inoculated branch.; (c) Control with inoculation wound (arrow) 20 d after inoculation. (d,e) Young sequoias inoculated with a pure culture of Neofusicoccum parvum 20 d after inoculation, with dieback of the whole twig above and below the inoculation point. Arrows indicate the inoculation point.

Figure 6.
Appearance of the trunk at inoculation point 4 months after inoculation. (a,b) Controls with healed wounds; (c,d) Young sequoias inoculated with a pure culture of Neofusicoccum parvum, with necrosis symptoms of the trunk.
Figure 6.
Appearance of the trunk at inoculation point 4 months after inoculation. (a,b) Controls with healed wounds; (c,d) Young sequoias inoculated with a pure culture of Neofusicoccum parvum, with necrosis symptoms of the trunk.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Identified Botryosphaeriaceae species isolated on Sequoiadendron giganteum trees in Canton of Geneva.
Table 1.
Identified Botryosphaeriaceae species isolated on Sequoiadendron giganteum trees in Canton of Geneva.
| Tree | Localisation | Disease Severity 1 | Sampled Material | Isolate Nb | Pathogen Species | GenBank Accession | Isolate Accession |
|---|---|---|---|---|---|---|---|
| No1 | Vésenaz | ++ | twig | C | N. parvum | NA | NA |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R1 | N. parvum | MH500829 | UASWS1756 |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R2 | N. parvum | MH500830 | UASWS1757 |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R3 | N. parvum | MH500831 | UASWS1758 |
| No2 | Geneva (Cimetière des Rois) | +++ | twig | R4 | N. parvum | MH500832 | UASWS1759 |
| No3 | Geneva (Parc Barton) | +++ | twig | B1 | N. parvum | MH500826 | UASWS1753 |
| No3 | Geneva (Parc Barton) | +++ | twig | B2 | N. parvum | MH500827 | UASWS1754 |
| No3 | Geneva (Parc Barton) | +++ | twig | B3 | N. parvum | MH500828 | UASWS1755 |
| No4 | Lullier | + | twig | A1 | Fusicoccum aesculi | MH500836 | UASWS1763 |
| No5 | Lullier | +/− | twig | B1 Lullier | Sordaria sp. | MH500837 | UASWS1764 |
| No6 | Lullier | + | twig | C1 | N. parvum | MH500838 | UASWS1765 |
| No6 | Lullier | bark | D1 | Diplodia mutila | MH500839 | UASWS1766 | |
| No7 | Chêne-Bourg | ++ | stroma on trunk | E1 | N. parvum | MH500840 | UASWS1767 |
| No8 | Carouge | + | twig | FV2 | Diplodia seriata | MH500841 | UASWS1768 |
| No8 | Carouge | + | twig | 1.1. | N. parvum | MH500822 | UASWS1749 |
| No8 | Carouge | + | twig | 2.1 | Diplodia mutila | MH500823 | UASWS1750 |
| No8 | Carouge | + | twig | 2.2 | Sordaria sp. | MH500824 | UASWS1751 |
| No8 | Carouge | + | twig | 3.1 | Phomopsis sp. | MH500825 | UASWS1752 |
| No9 | Geneva (Parc des Bastions) | ++ | twig | Bastions 1.3 | N. parvum | MH500833 | UASWS1760 |
| No9 | Geneva (Parc des Bastions) | ++ | twig | Bastions 3.1 | Fusicoccum aesculi | MH500834 | UASWS1761 |
| No9 | Geneva (Parc des Bastions) | ++ | twig | Bastions 4.1 | Fusicoccum aesculi | MH500835 | UASWS1762 |
1 arbitrary estimation of disease severity. +, ++, +++ and +/− mean different degrees of disease severity.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Haenzi, M.; Cochard, B.; Chablais, R.; Crovadore, J.; Lefort, F. Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland. Forests 2021, 12, 434. https://doi.org/10.3390/f12040434
AMA Style
Haenzi M, Cochard B, Chablais R, Crovadore J, Lefort F. Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland. Forests. 2021; 12(4):434. https://doi.org/10.3390/f12040434
Chicago/Turabian StyleHaenzi, Martine, Bastien Cochard, Romain Chablais, Julien Crovadore, and François Lefort. 2021. "Neofusicoccum parvum, A New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland" Forests 12, no. 4: 434. https://doi.org/10.3390/f12040434
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.