
Responses of Larix principis-rupprechtii Radial Growth to Climatic Factors at Different Elevations on Guancen Mountain, North-Central China


Abstract
:1. Introduction
2. Materials and Methods
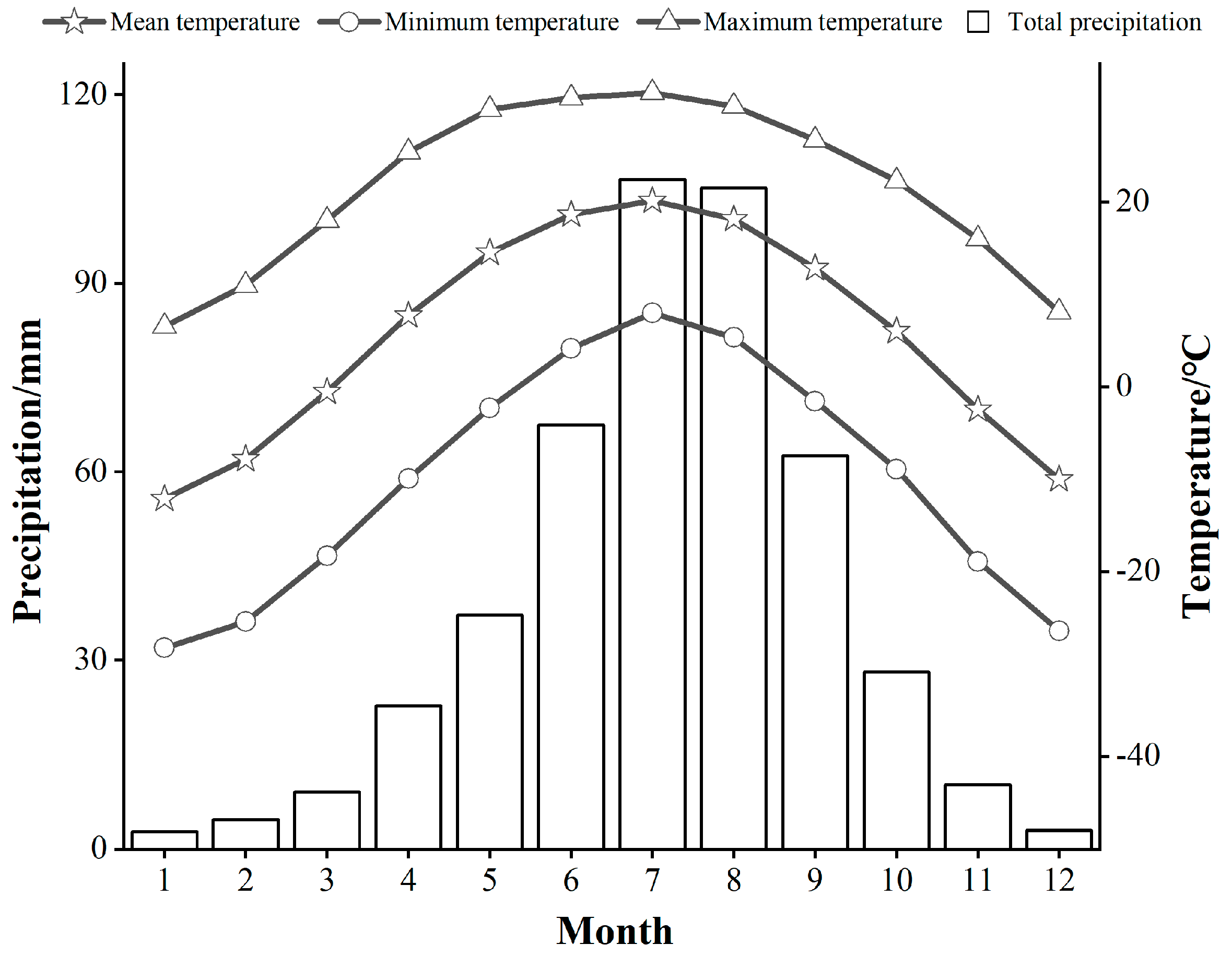
2.1. Overview of the Study Area
2.2. Sample Collection and Chronology Establishment
2.3. Meteorological Data Collection and Processing
2.4. Data Analysis
3. Results
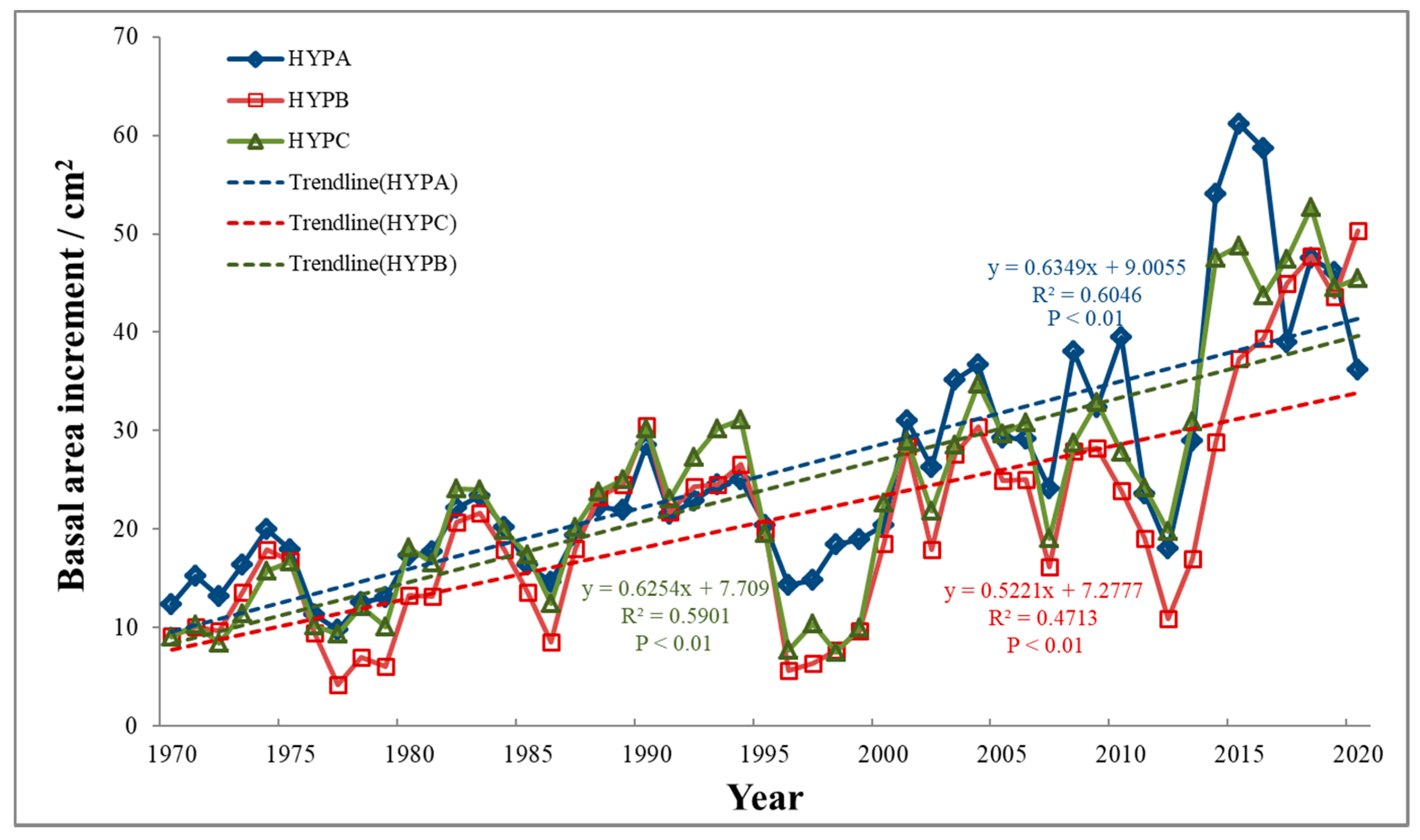
3.1. Chronological Characteristics Analysis
3.2. Correlation Analysis of the L. principis-rupprechtii Chronologies and Monthly Climatic Factors at Different Elevations
3.3. Redundancy Analysis of the L. principis-rupprechtii Chronologies and Climatic Factors at Different Elevations
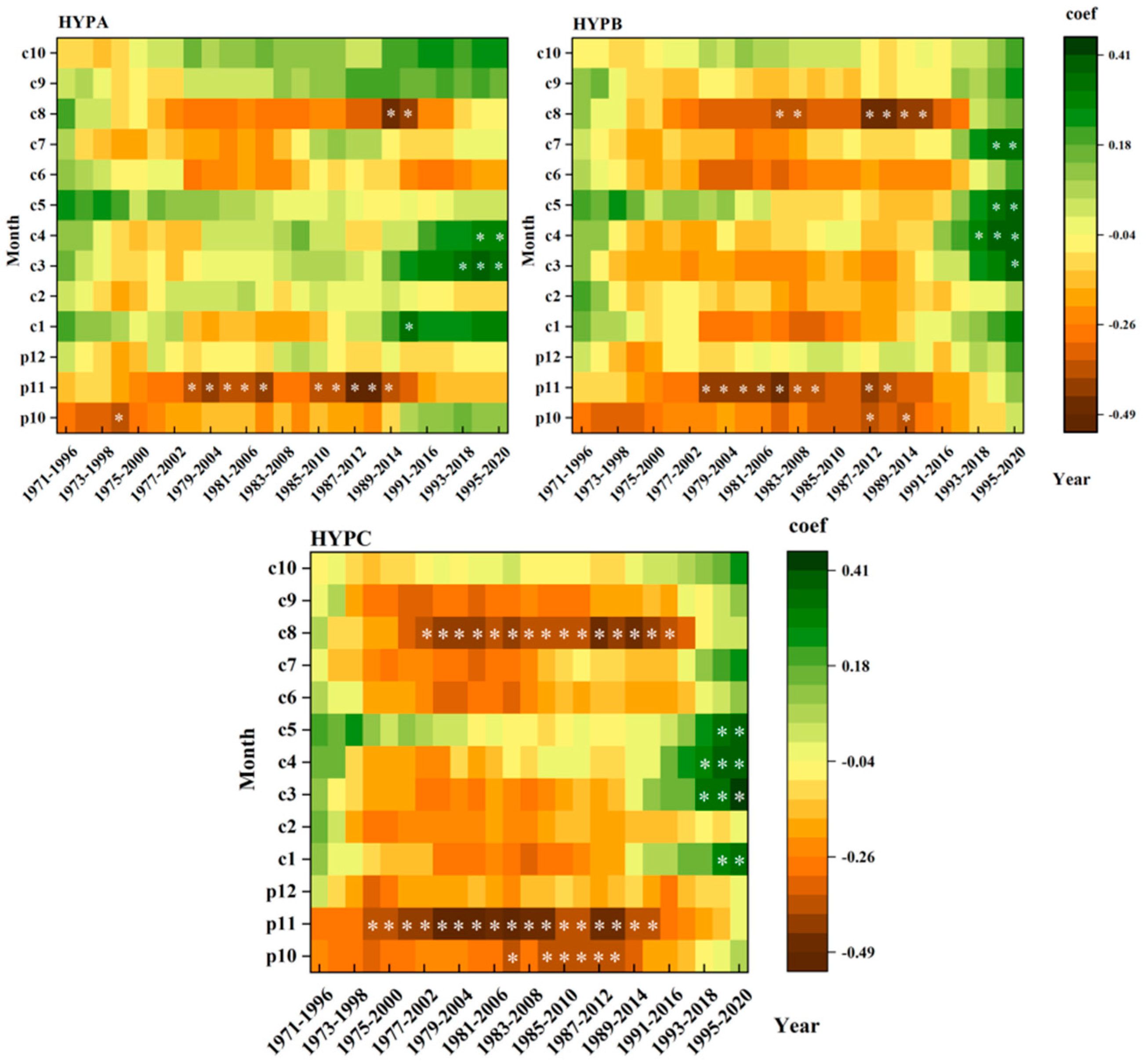
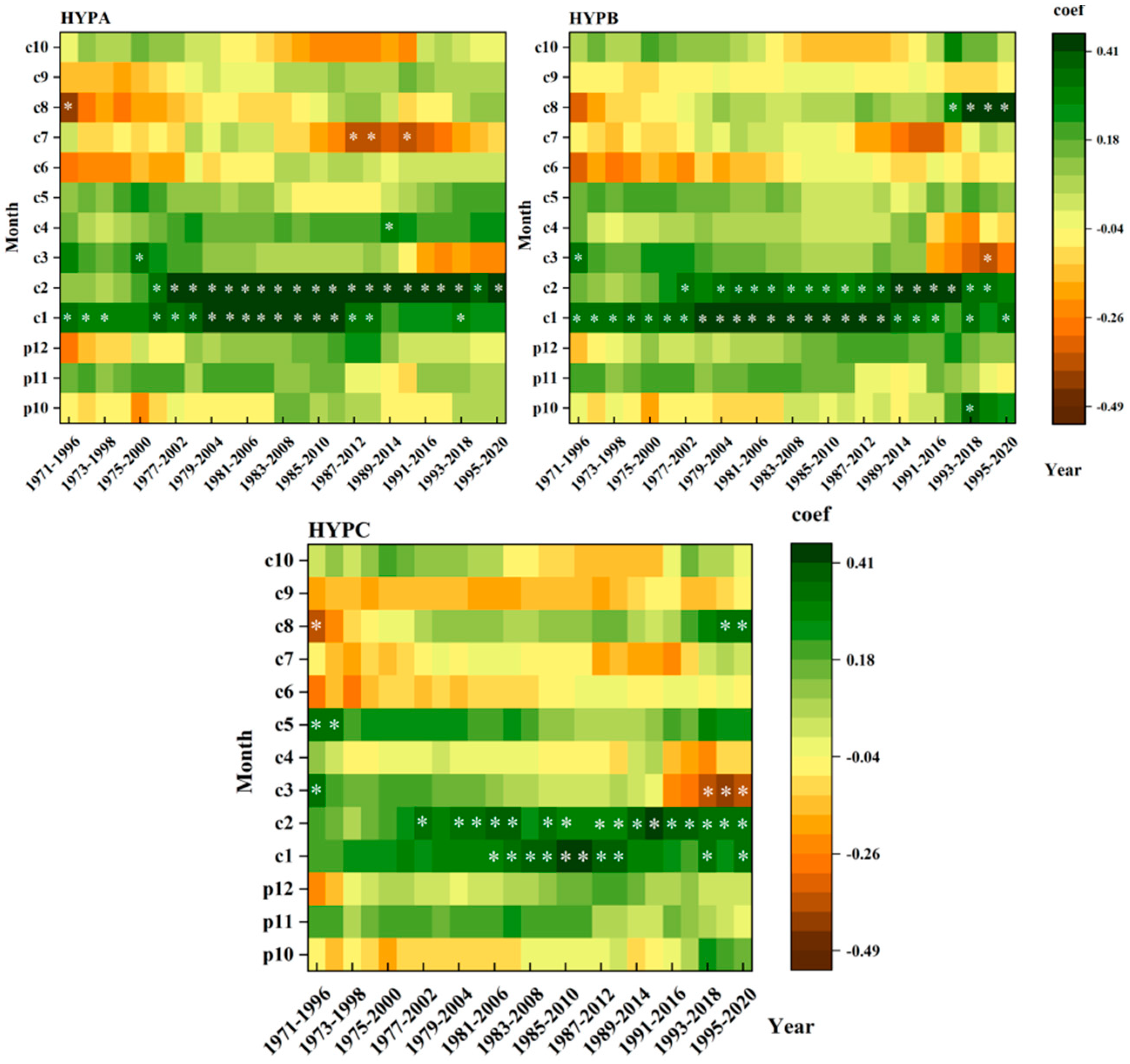
3.4. Dynamic Relationships between the L. principis-rupprechtii Chronologies and Climatic Factors at Different Elevations
4. Discussion
4.1. Differences in L. principis-rupprechtii Radial Growth at Different Elevations
4.2. Correlations between L. principis-rupprechtii Radial Growth and Climatic Factors at Different Elevations
4.3. Stability of L. principis-rupprechtii Radial Growth and Climatic Factors at Different Elevations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nowacki, G.J.; Abrams, M.D. The Demise of Fire and “Mesophication” of Forests in the Eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. Is climate an important driver of post-European vegetation change in the Eastern United States? Glob. Chang. Biol. 2015, 21, 314–334. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, A.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Engelbrecht, B.M.J. Forests on the brink: An analysis of the physiological vulnerability of different trees to drought shows that forests around the globe are at equally high risk of succumbing to increases in drought conditions. Nature 2012, 491, 675–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Shao, X. A Preliminary analysis on response of tree-ring density to climate in the Qinling Mountains of China. Chin. J. Appl. Ecol. 1994, 5, 253–256. [Google Scholar]
- Qin, J.; Bai, H.; Zhou, Q.; Wang, J.; Li, S.; Gan, Z.; Bao, G. Radial growth response of Abies fargesii to climate change from different elevations at timberline of Niubeiliang Natural Reserve. Arid Land Geogr. 2017, 40, 147–155. (In Chinese) [Google Scholar]
- Kang, Y.; Liu, J.; Dai, S.; He, X. Characteristics of ring-width chronologies of Larix chinensis and their responses to climate change at different elevations in Taibai Mountain. J. Northwest Univ. 2010, 38, 141–147. (In Chinese) [Google Scholar]
- Wu, X. Application of tree ring analysis to the study on environment variation. Quat. Sci. 1990, 10, 188–196. (In Chinese) [Google Scholar]
- Yang, Y.; Liu, Y.; Shi, J.; Cai, Q.; Song, J.; Yi, L.; Li, Q.; Sun, H. Experimental method of density for tree-ring research and Its application in research of tree ring data from Zhungeer County. Inn. Mongalia (In Chinese). 2006, 29, 639–645. (In Chinese) [Google Scholar]
- Christian, Z.; Franco, B. Dendroclimatic calibration in R: The bootRes package for response and correlation function analysis. Dendrochronologia 2013, 31, 68–74. [Google Scholar] [CrossRef]
- Preechamart, S.; Pumijumnong, N.; Payomrat, P.; Buajan, S. Variation in climate signals in Teak tree-ring chronologies in two different growth areas. Forests 2018, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Klesse, S.; von Arx, G.; Gossner, M.M.; Hug, C.; Rigling, A.; Queloz, V. Amplifying feedback loop between growth and wood anatomical characteristics of Fraxinus excelsior explains size-related susceptibility to ash dieback. Tree Physiol. 2021, 41, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shao, X.; Zhang, Y. Research progress on the response of radial growth to climatic factors at different altitudes. J. Earth Environ. 2012, 3, 845–854. (In Chinese) [Google Scholar]
- Yang, R.; Fan, Z.; Li, Z.; Wen, Q. Radial growth of Pinus yunnanensis at different elevations and their responses to climatic factors in the Yulong Snow Mountain, northwest Yunnan, China. Acta Ecol. Sin. 2018, 38, 8983–8991. (In Chinese) [Google Scholar]
- Zhang, Y.; Bergeron, Y.; Gao, L.; Zhao, X.; Wang, X.; Drobyshev, L. Tree growth and regeneration dynamics at a mountain ecotone on Changbai Mountain, northeastern China: Which factors control species distributions? Ecol. Sci. 2014, 21, 387–404. [Google Scholar] [CrossRef]
- Panthi, S.; Brauning, A.; Zhou, Z.; Fan, Z. Growth response of Abies georgei to climate increases with elevation in the central Hengduan Mountains, southwestern China. Dendrochronologia 2018, 47, 1–9. [Google Scholar] [CrossRef]
- Fan, Z.; Brauning, A.; Cao, K.; Zhu, S. Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, southwestern China. For. Ecol. Manag. 2009, 258, 306–313. [Google Scholar] [CrossRef]
- Chen, L.; Yin, Y.; Zhao, D.; Yuan, Q.; Wu, S. Climate response of tree growth along an altitudinal gradient in the Changbai Mountains, northeast China. Acta Ecol. Sin. 2014, 34, 1568–1574. (In Chinese) [Google Scholar]
- Oberhuber, W.; Kofler, W. Topographic influences on radial growth of Scots pine (Pinus sylvestris L.) at small spatial scales. Plant. Ecol. 2000, 146, 231–240. [Google Scholar] [CrossRef]
- Sang, W.; Wang, Y.; Su, H.; Lu, Z. Response of tree-ring width to rainfall gradient along the Tianshan Mountains of northwestern China. Chin. Sci. Bull. 2007, 52, 2954–2962. [Google Scholar] [CrossRef]
- Esper, J.; Frank, D.C.; Wilson, R.J.S.; Büntgen, U.; Treydte, K. Uniform growth trends among central Asian low- and high-elevation juniper tree sites. Trees 2007, 21, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Dario, M.B.; Nell, P. Convergence in drought stress, but a divergence of climatic drivers across a latitudinal gradient in a temperate broadleaf forest. J. Biogeogr. 2015, 42, 925–937. [Google Scholar] [CrossRef] [Green Version]
- D’Arrigo, R.; Wilson, L.; Liepert, B.; Cherubini, P. On the divergence problem’ in northern forests: A review of the tree-ring evidence and possible causes. Glob. Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Kurz-Besson, C.B.; Lousada, J.L.; Gaspar, M.J.; Correia, I.E.; David, T.S.; Soares, P.M.; Cardoso, R.M.; Russo, A.; Varino, F.; Mériaux, C.; et al. Effects of recent minimum temperature and water deficit increases on Pinus pinaster radial growth and wood density in southern Portugal. Front. Plant Sci. 2016, 7, 1170. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bai, H.; Ji, Q.; Li, M.; Ma, C.; Li, X.; Yang, C. Variations of plant community diversity with latitudinal gradients in Lvliang Mountains in Shanxi Province. Value Eng. 2016, 20, 198–201. (In Chinese) [Google Scholar]
- Qin, L.; She, L. Studies on the plant diversity in Guancen Mountains forest area. J. Taiyuan Norm. Univ. 2011, 10, 142–146. (In Chinese) [Google Scholar]
- Wang, Z.; Li, Q.; Liu, Y.; Yang, Y.; Ren, M.; Cui, L. PDSI variations recorded by tree rings in the northern Lvliang Mountains during the past 175 years. J. Earth Environ. 2020, 11, 72–80. (In Chinese) [Google Scholar]
- Pompa-García, M.; Cerano-Paredes, J.; Fuléa, P.Z. Variation in radial growth of Pinus cooperi in response to climatic signals across an elevational gradient. Dendrochronologia 2013, 31, 198–204. [Google Scholar] [CrossRef]
- Shrestha, K.B.; Chhetri, P.K.; Bista, R. Growth responses of Abies spectabilis to climate variations along an elevational gradient in Langtang National Park in the central Himalaya, Nepal. J. For. Res. 2017, 22, 274–281. [Google Scholar] [CrossRef]
- Babushkina, E.; Belokopytova, L.; Zhirnova, D.; Barabantsova, A.; Vaganov, E. Divergent growth trends and climatic response of Picea obovata along elevational gradient in Western Sayan mountains, Siberia. J. Mt. Sci. 2018, 15, 2378–2397. [Google Scholar] [CrossRef] [Green Version]
- Cai, Q.; Liu, Y.; Bao, G.; Lei, M.; Sun, B. Tree-ring-based May-July mean temperature history for Lvliang Mountains. Chin. Sci. Bull. 2010, 55, 2033–2039. (In Chinese) [Google Scholar]
- Li, Q.; Liu, Y.; Cai, Q.; Sun, J.; Yi, L.; Song, H.; Wang, L. Reconstruction of annual precipitation since 1688A.D. from Ningwu region, Shanxi Province. Quat. Sci. 2006, 26, 999–1006. (In Chinese) [Google Scholar]
- Yi, L.; Liu, Y.; Song, H.; Li, Q.; Cai, Q.; Yang, Y.; Sun, J. Summer temperature variations since 1676 AD in Luya Mountain, Shanxi province of China, inferred from tree rings. J. Glaciol. Geocryol. 2006, 28, 330–336. (In Chinese) [Google Scholar]
- Zhang, W.; Jing, Y.; Wang, M.; Zhang, L.; Dong, M.; Guo, Y. Responses of radial growth to climate warming in Picea meyeri trees growing at different elevations on the southern slope of Luya Mountain. Chin. J. Plant Ecol. 2013, 37, 1142–1152. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Wang, M.; Zhang, L.; Dong, M. Responses of radial growth in Larix principis-rupprechtii to climate change along an elevation gradient on the southern slope of Luya Mountain. Acta Ecol. Sin. 2015, 35, 6481–6488. (In Chinese) [Google Scholar]
- Li, Y.; Wang, S.; Niu, J.; Fang, K.; Li, X.; Li, Y.; Bu, W.; Li, Y. Climate-radial growth relationship of Larix principis-rupprechtii at different altitudes on Luya Mountain. Acta Ecol. Sin. 2016, 36, 1608–1618. (In Chinese) [Google Scholar]
- Hao, X.; Shangguan, T. Plant species diversity of the forestry community in Guancen Mountain at the source of Fen River of Shanxi. J. Anhui Agric. Sci. 2009, 37, 6232–6235. (In Chinese) [Google Scholar]
- Ma, L. Study on biodiversity characteristic in Guancen Mountain. Shaanxi For. Sci. Technol. 2014, 43, 19–21. (In Chinese) [Google Scholar]
- Xie, M. Investigation on typical vegetation communities and ancient and famous trees in Guancen mountain forest area. Inn. Mong. For. Investig. Des. 2015, 38, 90–93. (In Chinese) [Google Scholar]
- An, S. Vertical distribution law of forest vegetation and soil types in Guancen Mountai. Shaanxi For. Sci. Technol. 1998, 2, 20–22. (In Chinese) [Google Scholar]
- Zhai, D.; Zhang, T. Vegetation and its utilization in Guancen mountain. J. Shaanxi Norm. Univ. 1992, 4, 39–43. (In Chinese) [Google Scholar]
- Fang, K.; Yang, B.; Zheng, H. Research methods of dendrochronology and its application in global change. Quat. Sci. 2015, 35, 1283–1293. (In Chinese) [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Fritts, H. Tree Rings and Climate; Academic Press: London, UK, 1976; pp. 511–529. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–75. [Google Scholar]
- Holmes, R.; Adams, R.; Fritts, H. Tree-Ring Chronologies of Western North America: California, Eastern Oregon and Northern Great Basin. Chronology Series VI; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1986. [Google Scholar]
- Cook, S.P.; Hain, F.P. Defensive mechanisms of loblolly and shortleaf pine against attack by southern pine beetle, Dendroctonus frontalis Zimmermann, and its fungal associate, Ceratocystis minor (Hedgecock). Hunt. J. Chem. Ecol. 1986, 12, 1397–1406. [Google Scholar] [CrossRef]
- Wu, X. Tree Rings and Climate Change; Beijing Meteorological Press: Beijing, China, 1990. [Google Scholar]
- Chen, Y.; Chen, F.; Zhang, H.; Hu, M.; Wang, S. Strong link of large volcanic eruptions and climatic and hydrological changes recorded by tree rings in the river source area of Southern High Asia since 1200 A.D. Quat. Sci. 2021, 41, 323–333. (In Chinese) [Google Scholar]
- Liu, G.; Guo, A.; An, S.; Liu, W. Research progress in Palmer drought severity index and it’s application. J. Nat. Disaster 2004, 13, 21–27. (In Chinese) [Google Scholar]
- An, S.; Xing, J. Correction of Palmer drought severity model. Bull. Acad. Meteorol. Sci. 1986, 1, 75–82. (In Chinese) [Google Scholar]
- Wei, J.; Ma, Z. Comparison of palmer drought severity index, percentage of precipitation anomaly and surface humid index. Acta Geogr. Sin. 2003, 58, 117–124. (In Chinese) [Google Scholar]
- Braak, C. Canonical community ordination. Part I: Basic theory and linear methods. Ecol. Sci. 1994, 1, 127–140. [Google Scholar] [CrossRef]
- Olivier, T.; Masayoshi, N.; Zhao, H. Rare earth alkyl and hydride complexes bearing silylene-linked cyclopentadienyl-phosphido ligands. Synthesis, structures, and catalysis in olefin hydrosilylation and ethylene polymerization. Tetrahedron 2003, 59, 10525–10539. [Google Scholar] [CrossRef]
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Zhao, X.; Bai, X.; Li, J.; Wang, L.; Zhang, X.; Lu, X.; Chen, Z. Response of Larix olgensis at different elevations to climate change in the context of climate warming. Chin. J. Ecol. 2019, 38, 637–647. (In Chinese) [Google Scholar]
- Wigley, T.; Briffa, K.; Jones, P. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Climatol. Appl. Meteorol. 1984, 23, 201–213. Available online: https://www.jstor.org/stable/26181323 (accessed on 9 October 2021). [CrossRef]
- Han, Y.; Zhou, W.; Qi, L.; Zhou, L.; Zhong, Q.; Dai, L.; Yu, D. Tree radial growth-climate relationship in Changbai Mountain, Northeast China. J. Appl. Ecol. 2019, 30, 1513–1520. (In Chinese) [Google Scholar]
- Zhang, Y.; Yin, D.; Tian, K.; Zhang, W.; He, R.; He, W.; Sun, J.; Liu, Z. Radial growth responses of Picea likiangensis to climate variabilities at different altitudes in Yulong Snow Mountain, southwest China. Chin. J. Plant Ecol. 2018, 42, 629–639. (In Chinese) [Google Scholar]
- Yu, J.; Xu, Q.; Liu, C.; Luo, C.; Yang, J.; Li, J.; Liu, Q. Response of radial growth to climate change for Larix olgensis along an altitudinal gradient on the eastern slope of Changbai Mountain, northeast China. Chin. J. Plant Ecol. 2016, 40, 24–35. (In Chinese) [Google Scholar]
- Pan, H.L.; Li, M.H.; Cai, X.H.; Wu, J.; Du, Z.; Liu, X.L. Responses of growth and ecophsiology of plants to altitude. Ecol. Environ. Sci. 2009, 18, 722–730. [Google Scholar]
- Korner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef]
- Yang, B.; He, M.; Shishov, V.; Tychkov, L.; Vaganov, E.; Rossi, S.; Ljungqvist, C.; Bräuning, A.; Grießinger, J. New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proc. Natl. Acad. Sci. USA 2017, 114, 6966–6971. [Google Scholar] [CrossRef] [Green Version]
- Bai, T.; Deng, W.; Kuang, Y.; Liu, Y.; Ye, Q.; Niu, J.; Wen, L. Response of tree ring width in Cryptomeria japonica to climatic factors at different elevations in Lushan Mountain. Chin. J. Ecol. 2020, 39, 57–66. (In Chinese) [Google Scholar]
- Sun, Y.; Wang, L.; Chen, J.; Duan, J.; Shao, X.; Chen, K. Growth characteristics and response to climate change of Larix miller tree-ring in China. Sci. Chin. 2010, 40, 645–653. (In Chinese) [Google Scholar] [CrossRef]
- Hughes, M.K.; Kelly, P.M.; Pilcher, J.R. Climate from Tree Rings; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Li, J.; Yuan, Y. The role of snow in the growth of tree rings. J. Xinjiang Univ. 1984, 1, 85–92. (In Chinese) [Google Scholar]
- Begun, A.; Gerasimenyuk, N.; Korneev, A.; Molochkov, A.; Niemi, A. Gauge theory: Protein topology and dynamics. J. Bioenerg. Biomembr. 2018, 50, 500–501. [Google Scholar]
- Zhu, H.; Wang, L.; Shao, X.; Fang, X. Tree ring-width response of Picea schrenkiana to climate change. Geogr. Sin. 2004, 59, 863–870. (In Chinese) [Google Scholar]
- Gou, X.; Chen, F.; Jacoby, G.; Cook, E.; Yang, M.; Peng, J.; Zhang, Y. Rapid tree growth with respect to the last 400 years in response to climate warming, northeastern Tibetan Plateau. Int. J. Climatol. 2007, 27, 1497–1503. [Google Scholar] [CrossRef]
- Chen, C. Response of Radial Growth of Subalpine Coniferous Forest to Climate in Daofu County, Sichuan Province. Master’s Thesis, Chengdu University of Technology, Chengdu, China, 2019. [Google Scholar]
- Feng, X.H.; Cheng, R.M.; Xiao, F.W.; Wang, R.L.; Wang, X.R.; Gao, B.Q. The influence of temperature in north subtropical growing period on the radial growth of Masson Pine. J. Ecol. 2011, 30, 650–655. [Google Scholar]
- Cheng, R.; Feng, X.; Xiao, W.; Wang, R.; Wang, X.; Du, H. Response of net productivity of masson pine plantation to climate change in North Subtropical Region. Acta Ecol. Sin. 2011, 31, 2086–2095. (In Chinese) [Google Scholar]
- Zhang, T.; Yuan, Y.; Wei, W.; He, Q.; Zhang, R.; Yu, S.; Chen, F. Contrastive analysis and climate response of tree-ring width of Picea schrenkiana at the upper and lower forest limits in the middle section of the Kaiduhe River watershed. Arid Land Geogr. 2013, 36, 680–690. (In Chinese) [Google Scholar]
- Xue, R.; Jiao, L.; Liu, X.; Chen, K. Evaluation of the stability of the radial growth of Larix sibirica at different altitudes in response to climate change in Altai Mountains, Xinjiang. Chin. J. Ecol. 2021, 40, 1275–1284. (In Chinese) [Google Scholar] [CrossRef]
- Zhou, Z.; Jiang, Y.; Dong, M.; Tao, Y.; Wang, M.; Ding, X. Response of the relationship between radial growth and climatic factors to abrupt change of temperature along an altitudinal gradient on the northern slope of Changbai Mountain, northeast China. Acta Ecol. Sin. 2018, 38, 4668–4676. (In Chinese) [Google Scholar]
- Zhang, W.; Jiang, Y.; Dong, M.; Yang, Y.; Yang, H. Larix principis-rupprechtii’s radial growth-climate relationship at different elevations in Luya Mountains. J. Beijing Norm. Univ. 2011, 47, 304–309. (In Chinese) [Google Scholar]
- Wu, X.; Jiao, L.; Du, D.; Qi, C.; Xue, R. Comparison of the radial growth response of Picea crassifolia to climate change in different regions of the central and eastern Qilian Mountains. Forests 2021, 12, 1015. [Google Scholar] [CrossRef]
- Duan, J.; Wang, L.; Xu, Y.; Sun, Y.; Chen, J. Response of tree-ring width to climate change at different elevations on the east slope of Gongga Mountains. Geogr. Res. 2010, 29, 1940–1949. (In Chinese) [Google Scholar]
- Zhang, Y.; Yin, D.; Tian, K.; He, R.; He, M.; Li, Y.; Sun, D.; Zhang, W. Relationship between radial growth of Abies georgei and climate factors at different altitudes on the eastern slope of Yulong Snow Mountain, China. Chin. J. Appl. Ecol. 2018, 29, 2355–2361. (In Chinese) [Google Scholar]
- Jiang, Y.; Cao, Y.; Han, S.; Zhang, J.; Hao, L. Spatial variation and temporal instability in the growth/climate relationship of White Birch (Betula platyphylla Suk) in the Changbai Mountain, China. Forests 2021, 12, 589. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.; Wei, W.; Yu, S.; Zhao, Y.; Fan, Z.; Li, Y.; Zhang, R.; Zhang, T.; Shang, H. Variations of long-term palmer drought index in recent 354 years in Yili based on tree-ring record. Plateau Meteorol. 2011, 30, 355–362. (In Chinese) [Google Scholar]
- Qiao, J.; Wang, T.; Pan, L.; Sun, Y. Responses of radial growth to climate change in Pinus massoniana at different altitudes and slopes. Chin. J. Appl. Ecol. 2019, 30, 2231–2240. (In Chinese) [Google Scholar]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.-W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | Latitude | Longitude | Elevation (m) | Sample Size (Tree\Core) | Diameter at Breast Height (cm) * | Period |
|---|---|---|---|---|---|---|
| HYPA | 38°44′30″ N | 111°53′32″ E | 2303 | 24\48 | 43.73 ± 5.17 | 1899–2020 |
| HYPB | 38°43′46″ N | 111°52′46″ E | 2477 | 23\46 | 39.17 ± 3.55 | 1914–2020 |
| HYPC | 38°43′45″ N | 111°52′13″ E | 2539 | 23\46 | 42.25 ± 4.44 | 1919–2020 |
| Sampling Site | Elevation (m) | Slope Steepness (°) | Slope Aspect (°) | Soil | Canopy Availability | Air Pressure (kpa) |
|---|---|---|---|---|---|---|
| HYPA | 2303 | 27 | West by north 36 | Leached cinnamon, Brown earth | 0.6 | 77.36 |
| HYPB | 2477 | 13 | East by south 72 | Brown earth | 0.7 | 74.98 |
| HYPC | 2539 | 65 | West by north 5 | Brown earth | 0.4 | 74.42 |
| Sampling Site | HYPA | HYPB | HYPC |
|---|---|---|---|
| Common interval | 1930–2020 | 1950–2020 | 1950–2020 |
| SD | 0.397 | 0.351 | 0.305 |
| MS | 0.237 | 0.240 | 0.245 |
| r | 0.882 | 0.690 | 0.640 |
| AC1 | 0.875 | 0.684 | 0.634 |
| R1 | 0.376 | 0.634 | 0.638 |
| R2 | 0.350 | 0.650 | 0.748 |
| R3 | 0.385 | 0.631 | 0.596 |
| SNR | 5.427 | 64.163 | 35.217 |
| EPS | 0.844 | 0.985 | 0.972 |
| Sampling Site | HYPA | HYPB | HYPC |
|---|---|---|---|
| HYPA | 1 | - | - |
| HYPB | 0.666 ** | 1 | - |
| HYPC | 0.651 ** | 0.745 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, S.; Guo, Y.; Yang, Q.; Ren, R.; Han, Y. Responses of Larix principis-rupprechtii Radial Growth to Climatic Factors at Different Elevations on Guancen Mountain, North-Central China. Forests 2022, 13, 99. https://doi.org/10.3390/f13010099
Wang J, Li S, Guo Y, Yang Q, Ren R, Han Y. Responses of Larix principis-rupprechtii Radial Growth to Climatic Factors at Different Elevations on Guancen Mountain, North-Central China. Forests. 2022; 13(1):99. https://doi.org/10.3390/f13010099
Chicago/Turabian StyleWang, Jiachuan, Shuheng Li, Yili Guo, Qi Yang, Rui Ren, and Yijie Han. 2022. "Responses of Larix principis-rupprechtii Radial Growth to Climatic Factors at Different Elevations on Guancen Mountain, North-Central China" Forests 13, no. 1: 99. https://doi.org/10.3390/f13010099