
Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review
by
 , and
, and
Qinwang Xu
1,2,
Xuejiao Zhang
1,2,
Jiaxing Li
1,2,
Jiaru Ren
1,2,
Lili Ren
1,2,* and
Youqing Luo
1,2,* 1
Beijing Key Laboratory for Forest Pest Control, Beijing Forestry University, Beijing 100083, China
2
Sino-French Joint Laboratory for Invasive Forest Pests in Eurasia, Beijing Forestry University, Beijing 100083, China
*
Authors to whom correspondence should be addressed.
Forests 2023, 14(2), 174; https://doi.org/10.3390/f14020174
Submission received: 19 December 2022
/
Revised: 11 January 2023
/
Accepted: 16 January 2023
/
Published: 18 January 2023
(This article belongs to the Special Issue Forests Research in Beijing Forestry University: Commemorating the University’s 70th Anniversary)
Abstract
:Pine wilt disease (PWD) caused by the pine wood nematode (PWN) is China’s most serious forest disease in recent decades. Its complex disease system with unclear pathogenesis and differentiated pathogenicity makes it challenging to manage and control. PWD has spread rapidly in Northeast and Northwest China in recent years, exceeding previous predictions and the traditional understanding of its potential distribution. This paper reviewed the comprehensive risk of PWD in Northeast and Northwest China through the occurrence of influencing factors. Regarding PWN’s temperature resistance, all regions of China are suitable for PWD. In terms of altitude, the PWD system is mainly distributed below 1000 m. In China, Pinus and Larix species are confirmed host trees of PWN, while Monochamus alternatus and M. saltuarius are confirmed vector insects of PWN. In this case, PWD may occur in all low-altitude pine forests in China. Northeast China has a higher risk than Northwest China due to its distribution of suitable host trees and vector beetles at low altitudes. To confront the threat, two scientific bottlenecks are required to be demonstrated. The first is the pathogenicity of Chinese PWN strains to Chinese Picea and Abies species. The other is the geographical distribution and highest altitude distribution of other Monochamus species in the Palaearctic region of China and their PWN transmission ability. We also suggest strengthening plant quarantining in Northeast China. In conclusion, this review comprehensively analyzes the risk of PWD in Northeast and Northwest China in terms of temperature, altitude, host trees, and vector insects. It provides a new perspective and reference for the management and research of PWD in China.
1. Introduction
The pine wood nematode (abbreviated as PWN), Bursapherenchus xylophilus Steiner & Buhrer, 1934 (Nematoda: Rhabditida: Aphelenchoididae) is native to North America and was originally recorded in the United States, Canada, and Mexico [1,2,3]. It was first found in Pinus palustris in its origin in 1929 and then was named in 1934 [1]. Its first invasion was speculated to occur in Japan in 1905 before it had a name [4]. PWN has an invasion history of more than 100 years, with Asia and Europe as the main invasion areas, including Japan, China, the Republic of Korea, Portugal, and Spain [4,5,6,7,8]. PWN causes severe pine wilt disease (abbreviated as PWD) in the invasion area, a fatal disease of pine trees [5]. PWN invaded Taiwan and Hong Kong in China in the 1970s and the Chinese mainland in 1982 [9,10,11]. PWD is the most severe forest disease in China (hereafter, the Chinese mainland is referred to as China) in recent decades [12]. It has caused disastrous economic and ecological losses in the 40 years of invasion in China [12]. The associated economic losses in China reached CNY 19.5 billion in 2017, with an average annual loss of CNY 7.17 billion from 1998 to 2017 [13,14].
Generally, the 10 °C isotherms of annual average temperature were considered the distribution limits of PWD in China, and PWD would not occur north of the isotherms [15]. However, the distribution of PWD crossed the 10 °C isotherms and entered Northeast China in recent years, shattering this traditional understanding [16]. PWN increased in the new host tree genus Larix and new vector insect species Monochamus saltuarius in Northeast China, bringing a considerable threat to the pine forest resources there [17,18,19]. The current distribution of PWN in China also exceeded the potential geographical distribution areas predicted based on different climatic conditions (current climatic conditions, cold conditions, climate warming, and different warming modes) and various methods and models [20,21,22,23,24,25,26,27]. Those models and methods only considered climate factors and historical distribution data and did not consider PWD’s complex disease system, resulting in a considerable difference between prediction results and actual occurrence. According to Chinese national forest pests survey results and expert evaluation, PWD showed significant spreading trends in Northeast and Northwest China [28]. A more comprehensive approach is needed to analyze the risks of PWD in Northeast and Northwest China.
The risk of PWD in Northeast and Northwest China was reviewed in this paper, with the following purposes: (1) to study PWD’s invasion history in China and identify its spread trends in Northeast and Northwest China; (2) to analyze the risk of PWD in Northeast and Northwest China according to the occurrence factors of PWD based on the complexity of the PWD plant disease system and information on its occurrence and damage in China as well as in other invasion and origin countries; (3) to address problems that need to be urgently solved due to trends of spread and provide a reference for relevant research and management.
2. Trends of the Spread of PWD in Northeast and Northwest China
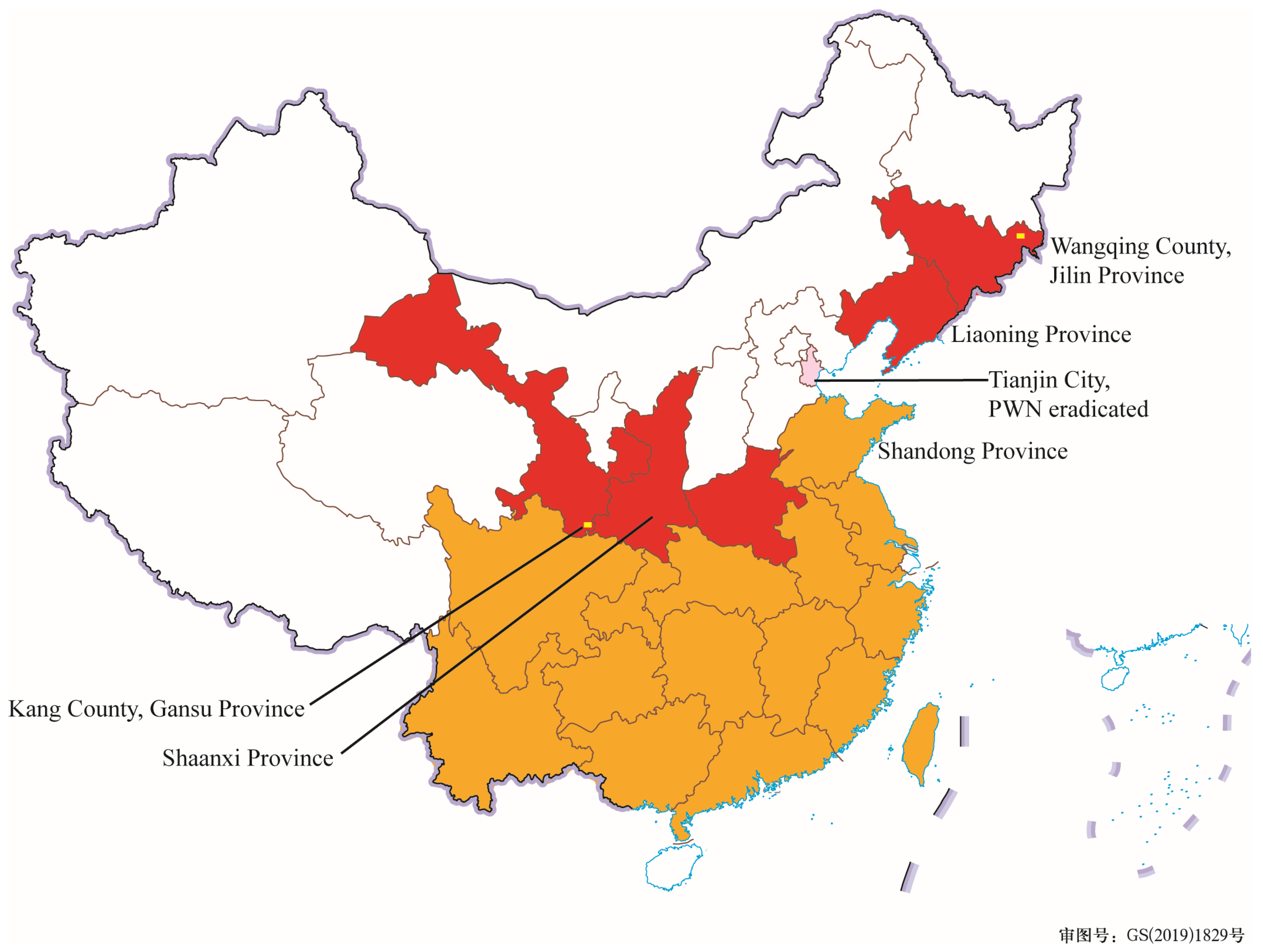
According to the announcement of the China National Forestry and Grassland Administration, there are 19 provincial epidemic areas of PWD in the Chinese mainland in 2022: Anhui Province, Chongqing City, Fujian Province, Gansu Province, Guangdong Province, Guangxi Zhuang Autonomous Region, Guizhou Province, Henan Province, Hubei Province, Hunan Province, Jiangsu Province, Jiangxi Province, Jilin Province, Liaoning Province, Shandong Province, Shaanxi Province, Sichuan Province, Yunnan Province, and Zhejiang Province, as shown in Figure 1. PWD occurred in all seven administrative geographic regions (Northeast China, East China, North China, Central China, South China, Southwest China, and Northwest China). Only the epidemic areas in North China (Tianjin City) were eradicated. The provincial epidemic areas in Northeast China included Liaoning Province and Jilin Province, and the provincial epidemic areas in Northwest China included Shaanxi Province and Gansu Province. Currently, the PWD’s most frontier epidemic points in Northeast and Northwest China are Wangqing County, Yanbian Korean Autonomous Prefecture, Jilin Province, and Kang County, Longnan City, Gansu Province, respectively.
The spread of PWD in China has passed through two periods [29,30]. The first period involved PWD spreading westward and northward from Jiangsu Province and Guangdong Province (introduced from Hong Kong) as the center from 1982 to 2008 [29,30,31]. During this period, the epidemic area was generally distributed in southern China, and only Qingdao, Yantai, and Weihai (three cities in the coastal areas of Shandong Peninsula) in Shandong Province, were the epidemic areas in northern China (the orange-filled areas in Figure 1). In the second period, from 2009 until the present, PWD continued invading more northern provinces (the red- and pink-filled areas in Figure 1). In 2009, PWD entered Shaanxi Province and Northwest China for the first time. In 2016, PWD entered Liaoning Province and Northeast China for the first time by a jump, crossing North China. PWD invaded Jilin Province in Northeast China and Gansu Province in Northwest China in 2021, showing noticeable spreading trends to the northeast and northwest.
3. The Complexity of PWD
The genus Bursapherenchus includes more than 70 species, mainly distributed in the northern hemisphere [32,33]. Only the PWN B. xylophilus and the red ring nematode B. cocophilus have significant plant pathogenicity in the genus Bursapherenchus, which are generally recognized as agricultural and forestry pests of global importance [33,34]. We cannot gain management experience from relative pests because PWN is a pest of conifers, while B. cocophilus is a pest of Palmae. PWD is difficult to manage due to its complex disease system composition, pathogenicity differentiation, and unclear pathogenesis. It is also challenging to copy other countries’ experiences and control strategies directly.
3.1. Complex Plant Disease System
The occurrence of PWD requires four conditions simultaneously: suitable environmental conditions, the PWN Bursapherenchus xylophilus, host pine trees, and vector insects (Monochamus species).
PWN has two developmental forms in its life cycle, the propagative form (egg; first-, second-, third-, and fourth-stage propagative juveniles; and adult males and females) and the dispersal form (third- and fourth-stage dispersal juveniles) [35]. The PWN propagative form feeds on parenchymatous cells of resin canals and fungi in pine trees. The dispersal third-stage juveniles of PWN (JIIIs) gather around the pupa chamber of Monochamus in winter. Before the emergence of the adult Monochamus, the JIIIs transform into dispersal fourth-stage juveniles (JIVs) and enter the Monochamus body [35]. The life cycles of PWN and Monochamus are coordinated by chemical signals [36,37]. The dispersal JIIIs secrete ascarosides that promote Monochamus alternatus pupation, and M. alternatus adults secrete ascarosides that attract the dispersal JIVs [37]. After eclosion, the Monochamus beetles migrate to tender branches of healthy pine trees for feeding and weakened pine trees for oviposition. The nematodes escape from Monochamus and enter pine trees through the wounds caused by the beetles, completing the transmission between trees. The infected pine generally shows wilting symptoms after a minimum of 3–4 weeks of infection.
In addition, the microorganisms (bacteria and fungi) associated with PWN and Monochamus participate in the development and pathogenicity of PWD [36,38,39,40,41]. Some endophytic microorganisms (bacteria and nematodes) in pine trees play a protective role [41,42]. These species constitute a complex PWD plant disease system.
The vector insect Monochamus species usually fly short distances in the wild when living in suitable environments, although they have the ability to fly a considerable distance [43,44,45,46]. The long-distance transmission of PWD mainly occurs via human activities such as the transportation of infested trees and wood. The complex disease system, coupled with human activities, has led to the complexity of PWD occurring on a spatiotemporal scale.
3.2. Pathogenic Differentiation in Different Regions
The difference in the biological composition of the PWD system has led to the difference in occurrence and damage. The pathogenicity of PWN strains, the species and pathogenicity of associated bacteria of PWN, the transmission capacity of different Monochamus species, and the resistance of different host tree species vary significantly in different regions.
According to host tree mortality and nematode reproduction, Li classified different geographic strains of PWN into three categories [47]. They were high-pathogenicity strains in China and Portugal, moderate-pathogenicity strains in Japan, and low-pathogenicity strains in Canada [47]. The PWN strains from Japan and Canada showed different pathogenicity to Pinus sylvestris, Larix occidentalis, and Picea mariana [48]. Even different geographic populations of nematodes in the same country had different pathogenicity [49]. The pathogenicity of the surface bacteria of PWN in America was weaker than in China [50]. The Japanese PWN strain can cause the death of Cedrus deodara, while the Chinese strain does not cause harm because of the difference in species and pathogenicity of the bacteria on the PWN surface [51,52,53]. The number of PWNs carried by Monochamus in North America is lower than that carried by the Asian species [30]. Due to long-term coevolution, the PWD system does not harm native tree species, while most non-native pine trees are susceptible [5]. The above differences lead to the stronger pathogenicity of the PWD system in East Asia than in the area of origin.
3.3. Unclear Pathogenesis
The pathogenesis of PWD is complex, and there are many pathogenicity hypotheses, such as phytotoxin, cellulase, terpenoid, PWN, and bacteria hypotheses [54,55]. It is widely believed that PWNs and bacteria create phytotoxins and cellulases, which result in pine tree defense, the production of terpenoids, and the formation of cavities that impede water transport, and finally, pine tree wilting [54,56]. However, the critical pathogenic factor of PWD is still unknown. In recent years, research on the functional genes of PWNs has developed rapidly, and some essential genes have been identified using RNAi and other new technologies [57,58].
4. PWN and Influencing Environmental Conditions
4.1. Temperature
Previous statistical studies showed that PWNs only caused damage where the average summer temperature was higher than 20 °C in North America and Japan [59]. PWD did not occur in areas with an annual average temperature below 10 °C in Japan [30]. Areas north of the 10 °C isotherms of the annual average temperature were also generally considered unsuitable for the survival of PWN in China [15]. PWD invaded Liaoning Province in 2018 and Jilin Province in 2021, breaking the traditional understanding. The average annual temperature in Wangqing County, Jilin Province, is only 3.8–4.7 °C, far lower than 10 °C [60]. Wangqing County is the northernmost occurrence point in China at present.
The nematode is a group of organisms with high tolerance to low temperatures and highly stressing conditions. Panagrolaimus davidi survival in the Antarctic habitat is the best example of an animal surviving intracellular freezing [61,62]. Viable soil nematodes were isolated in Pleistocene permafrost deposits of the Kolyma River Lowland with a natural cryopreservation of 30,000–40,000 years [63]. The common model nematode species, Caenorhabditis elegans, can be stored in liquid nitrogen (−196 °C), and 35–45% of animals can be revitalized after being frozen [64]. Caenorhabditis elegans survived the atmospheric breakup of Space Shuttle Columbia [65].
PWN’s ancestor originated from eastern Eurasia’s cold region [66]. The survival rates of propagative and dispersal juveniles of PWN were stable at about 95% in the gradient temperature change to −15 °C [67]. The PWN dispersal JIIIs can resist low-temperature stress by cryptobiosis, and their average survival rates were 1.7% after −80 °C treatment for 30 days and 82.2% after −20 °C treatment for 30 days [68]. The PWN population in southern China had a solid cold tolerance, indicating that the population invading from southern China to northern China could successfully invade without adaptation [69]. Sequencing analysis of the whole genome of PWNs from different geographic populations in China showed that the geographic-related SNPs were mainly located in the GPCR gene family related to adaptation, which indicated that nematodes had been evolving to adapt to different temperatures [31].
The above evidence proves that PWNs have the ability and genetic basis for resisting low temperatures. Therefore, PWN can survive in all regions of China, regardless of temperature [70]. Moreover, the temperature is not even a limiting factor for the spread of PWN in Northeast and Northwest China.
4.2. Altitude
Altitude is an essential limiting factor in the occurrence of PWN. With the increase in altitude, temperature and precipitation change, affecting the distribution of Pinaceae and Monochamus species, and consequently affecting the distribution of PWN.
The original distribution of PWN in the United States was mainly limited to states with an average altitude lower than 1000 m, according to scientific literature, the USDA nematode database, and USDA forest health annual highlights for each state [1,71,72,73]. The distribution records in some high-altitude states were unintentionally artificially introduced, such as Pinus nigra in Colorado [74]. The highest distribution of PWN in the United States is about 1500 m, isolated from the Rocky Mountains (Jane E. Stewart, personal communication, 13 September 2022). In general, B. xylophilus only harms introduced alien pine trees in the places of origin. Therefore, their relevant data are not as numerous and accurate in invasion countries.
For Japan, an invaded country, the highest distribution of PWNs is 1150 m in the northern region of Fuji Mountain, and the distribution limit of the vector insect Monochamus alternatus is about 1150 m [75]. PWD mainly occurs below 750 m in Japan [75]. The highest distribution of the vector insect Monochamus galloprovinciali is 1590 m in Europe [76]. Historical occurrences of PWD in China showed that the areas below 400 m in altitude had a serious infection, 400–700 m had a moderate infection, 700–1000 m had a sporadic infection, and those higher than 1000 m were non-occurrence areas [34,77]. Only one occurrence was reported at an altitude higher than 1000 m in China, at 1100 m in the southern region of the Qinling Mountains, Shaanxi Province, in 2009 [30].
To summarize, PWD mainly occurred in areas below 1000 m in altitude, although there are a few reports of findings above 1000 m. The terrain of China is divided into three steps from west to east, and the altitude decreases in turn. The distribution of PWD in China is limited to the second- and third-step regions, with an average altitude lower than 2000 m. The average altitude of the first-step Qinghai–Tibet Plateau is higher than 4000 m, which can be considered to have a low risk of PWD occurrence.
4.3. Other Environmental Conditions
Other environmental factors, such as precipitation and extreme abnormal weather, can also affect the occurrence of PWD, mainly by affecting the growth, vitality, and resistance of pine trees [30].
5. PWN and Its Host Trees
5.1. PWN Host Trees and Non-Host Trees in China
There are 11 genera in Pinaceae, all of them are distributed in China: Abies, Cathaya, Cedrus, Keteleeria, Larix, Nothotsuga, Picea, Pinus, Pseudotsuga, Pseudolarix, and Tsuga [78].
Pinus and Larix are natural host trees of PWNs in China [18]. PWNs can naturally infect at least 17 species in China [79]. They are Pinus armandii, P. bungeana, P. densiflora, P. elliottii, P. kesiya, P. koraiensis, P. luchuensis, P. massoniana, P. sylvestris var. mongolica, P. tabuliformis, P. taeda, P. taiwanensis, P. thunbergia, P. yunnanensis, Larix gmelinii var. principis-rupprechtii, L. kaempferi, and L. olgensis [79].
The genus Cedrus was proven to not be infected by PWD in China [51,80]. The genus Cathaya is an endemic and single-species genus of China distributed at an altitude above 1400 m in Guangxi and Sichuan Province [81]. The genus Keteleeria is endemic to China and Vietnam and distributed in southern China [81]. The genus Pseudolarix is an endemic and single-species genus in China distributed in the middle and lower reaches of the Yangtze River [81]. The genus Pseudotsuga is an endemic and single-species genus distributed in southern China [81]. These four genera are distributed in the epidemic provinces of PWD in China. Nevertheless, there has been no record of PWD infection or PWNs isolated from them. There have been no reports on the damage of PWNs to these four genera of trees worldwide. Therefore, they are cataloged as non-host species of PWN in China.
There has been no report of PWN-infested Abies, Picea, Pseudotsuga, and Tsuga in China. PWNs were isolated from Pseudotsuga menziesii in the United States and Canada [2,33]. There were also cases of artificially inoculated and dead Pseudotsuga menziesii, Tsuga mertensiana, and Tsuga heterophylla in Canada [82,83]. Pseudotsuga and Tsuga are mainly distributed in PWD-epidemic provinces, with no report of infection in China. Their risk of victimization in China is considered relatively low based on the historical evidence. Picea and Abies species are widely distributed in China and are important tree species in Northeast and Northwest China, with relatively high risks. As shown in Figure 2, the relationship between the genera of Pinaceae and PWNs in China can be divided into four categories.
The infection capacity of PWNs to Picea and Abies is unclear in China. In terms of the history of its occurrence, PWNs have invaded China for 40 years, spreading through over 19 provinces, with no report of PWNs infesting spruce and fir. Meanwhile, PWNs have an invasion history of more than 100 years, with no report that PWNs having killed spruce or fir forests on a large scale worldwide.
There are only a few reports on isolation from field trees, as shown in Table 1. PWNs were isolated from spruce and fir trees at its origin [2,5,33,84,85]. In the invasion area, PWNs were only isolated from fir and spruce in Japan [4,5]. However, the damage to non-Pinus hosts in Japan was almost negligible [4].
Additionally, there are only a few reports on death caused by artificial inoculation for Picea and Abies seedlings, as shown in Table 1. Artificial inoculation experiments in different areas showed the pathogenicity difference of PWN to spruce and fir. In general, artificial inoculation with PWNs caused lower mortality of Picea and Abies trees than Pinus [82,83]. The pathogenicity of PWN strains to Picea mariana in different regions was different [48]. Inoculation studies also showed that Picea abies was not susceptible to PWNs [87]. Nunes et al. found that PWNs were confined to the inoculation site, and the number of nematodes decreased with the extension of the inoculation time [87]. In addition, almost all the existing PWN inoculation experiments were conducted with seedlings. It is difficult to artificially inoculate grown-up trees due to the slow rate of disease progression and death.
All these factors indicate a complex relationship between PWN strains and Picea and Abies species in different regions. Thus, it is necessary to inoculate Chinese Picea and Abies trees with Chinese PWN strains to test their pathogenicity due to the differentiation of PWN pathogenicity and tree resistance in different regions.
5.2. Pinaceae in Northeast and Northwest China
The coniferous forests in Northeast China mainly include the Larix gmelinii forest in Greater Khingan, the broad-leaved and Pinus koraiensis mixed forest in Lesser Khingan, and the Pinus koraiensis and broad-leaved mixed forest in Changbai Mountain. The coniferous forests in Northwest China mainly include the coniferous forests in Qilian Mountains, the coniferous forests in Tian Shan, and the coniferous forests in Altai Mountains [88,89].
The main conifers in Northeast China are Pinus densiflora var. ussuriensis, P. koraiensis, P. pumila, P. sylvestris var. mongolica, P. sylvestris var. sylvestriformis, Larix gmelinii, L. olgensis, Picea jezoensis, Picea koraiensis, Abies holophylla, and Abies nephrolepis [88,89]. The main conifers in Northwest China are Pinus sibirica, Larix sibirica, Picea crassifolia, Picea obovate, Picea schrenkiana, and Abies sibirica [88,89]. The tree species with the lowest distribution altitude below 1000 m in Northeast and Northwest China are shown in Table 2, and they are all distributed in Northeast China [89].
Therefore, in terms of the host tree species and altitude distribution, the coniferous forests in Northeast China are much more threatened by PWD than those in Northwest China. Moreover, considering the rapid-spread of PWD in Northeast China and the fact that Picea and Abies are distributed in the low-altitude areas of Northeast China, it is urgent to use artificial inoculation methods to determine the pathogenicity of Chinese PWD strains to these species.
6. PWN and Its Vector Insects
6.1. Vector Insects of PWNs Worldwide
Many insects can carry PWNs, including Cerambycidae, Curculionidae, Buprestidae, Elateridae, and Termitidae [46]. Only the insects whose life history coincides with PWNs and that cause wounds to pine trees can become the vector insects of PWNs. All PWN vector insects have been from the genus Monochamus [43]. Monochamus comprises more than 100 species globally [90]. The Monochamus species originated from Africa and are now widely distributed in Eurasia, Africa, and North America [91]. Non-coniferous trees are the primary hosts of Monochamus in Africa, and conifers and non-coniferous trees are hosts in other distribution regions [92].
6.2. Monochamus Species in Northeast and Northwest China
Seven Monochamus species feed on conifers in China, as shown in Table 3 [93,94,95,96,97,98,99,100]. Furthermore, five species are distributed in Northeast China, while four live in Northwest China. Monochamus are distributed in most of Northeast and Northwest China.
Monochamus alternatus and M. saltuarius are confirmed vector insects of PWNs in China [17,19]. The former is the vector in southern China, and the latter is the primary vector in Northeast China, especially in Liaoning and Jilin provinces. The northernmost and westernmost distributions of M. alternatus are in Jilin Province and Shaanxi Province, respectively [93,101]. M. saltuarius is widely distributed in Northeast and Northwest China [93].
Monochamus galloprovincialis has been confirmed to be the vector of PWNs in Europe [43]. M. sutor can carry Bursapherenchus mucronatus and is considered a potential vector insect species of PWNs in Europe [102,103].
Therefore, vector insects are not obstacles to PWD in Northeast and Northwest China. The question is whether Monochamus galloprovincialis, M. sartor urussovii, and M. sutor can carry and spread PWNs in China and their transmission efficiency. Their highest altitude distribution in China is also unknown and needs to be studied.
Furthermore, Monochamus species (non-Chinese) have been the most intercepted longhorn beetles in the quarantine of imported plants in China in the recent twenty years [104]. Most of them have been found in imported timber from Europe and the United States [104]. Therefore, we also need to be alert to the introduction of Monochamus from abroad.
7. Comprehensive Risk of PWD in Northeast and Northwest China
7.1. Comprehensive Risk Analysis
Previous risk-analysis methods for PWD in China have focused on climate factors and historical distribution data without considering the complexity of the PWD system. The actual occurrence far exceeds all predictions and understandings of potential distribution. Thus, it is necessary to analyze the influencing occurrence factors of PWD one by one.
Our comprehensive risk analysis shows that the prevalence of PWD may occur in all low-altitude pine forests in China. PWNs have high adaptability to low temperatures, so it is speculated that it can adapt to various temperature zones in China. There are only a few reports of PWD in areas above 1000 m globally due to the limitation of high altitudes on the distribution of host pine trees and vector Monochamus. Therefore, the occurrence risk of PWD in the Qinghai–Tibet Plateau of China should be extremely low. From the perspective of biological factors of the PWD disease system, the Northeast region has a much higher risk compared with Northwest China due to its host tree and vector beetle distribution at lower altitudes.
7.2. Scientific Bottlenecks to Be Proven
7.2.1. The Pathogenicity of PWNs to Picea and Abies
Some Picea and Abies species in Northeast China are distributed below 1000 m, and thus face a severe risk for infection. Whether PWNs can cause spruce and fir death in China is a crucial scientific problem that needs to be solved first. Chinese PWN strains should be used to inoculate Chinese spruce and fir trees to observe pathogenicity due to PWN’s pathogenic differentiation. Validation tests of artificial inoculation for different species of Picea and Abies at different ages are suggested. Moreover, we also suggest isolating spruce and fir trees from the PWN epidemic area in Northeast China to see if PWNs have infected them but with no wilting symptoms.
If the PWN strains in China can successfully infest and kill spruce and fir tree species in China, Northeast China will face extremely severe risks. The spruce and fir trees in Northwest China would also be under threat. On the contrary, the risks and pressures faced by Picea and Abies species in Northeast and Northwest China would be significantly reduced.
7.2.2. The Distribution and PWN Transmissibility of Monochamus
The distribution information of five Monochamus species in Northeast and Northwest China needs to be investigated and confirmed. The Monochamus species may spread due to afforestation activities in recent decades. Moreover, the current literature data are relatively old. Studying the distribution of Monochamus species at different altitudes is also necessary, which is of great significance to understanding the altitude distribution limit of PWD in China. In addition, research on the transmission capacity of PWNs should also be carried out for Monochamus species in the Palaearctic region of China: M. galloprovincialis, M. sutor, and M. sartor urussovii.
7.3. Future PWD’s Management in Northeast and Northwest China
PWD has notable spreading trends in Northeast and Northwest China. PWD may occur in all low-altitude pine forests in China, and Northeast China faces a much higher risk than Northwest China.
Wood quarantine, epidemic monitoring, and eradication are the core measures for PWD control in China [12]. A total of 9 in 36 and 4 in 52 municipal administrative divisions in Northeast China and Northwest China, respectively, are epidemic areas of PWD. It is suggested that quarantine and prevention should be the first measure for most Northeast and Northwest China areas.
The long-distance spread of PWNs is mainly realized through the transportation of diseased wood. In 2021, living PWNs were detected in the pines imported from the United States by a company in Heilongjiang Province. Thus, from this aspect, the prevention of PWNs in Northeast China still needs to be strengthened. More effective quarantine measures should be set up for the high-risk non-epidemic areas in Northeast China, such as rapid detection and improving the traceability of infected wood.
Author Contributions
Conceptualization, Y.L.; methodology, Q.X., Y.L. and L.R.; investigation, Q.X., X.Z., J.L. and J.R.; writing—original draft preparation, Q.X.; writing—review and editing, Q.X., L.R., X.Z., J.L. and J.R.; visualization, Q.X.; supervision, Y.L. and L.R.; project administration, Y.L.; funding acquisition, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China, grant number 2021YFD1400900.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable.
Acknowledgments
We thank Jane E. Stewart from Colorado State University for providing information on the pine wood nematode in the United States. We thank the FAO (Food and Agriculture Organization of the United Nations) for its Forest Invasive Species Dgroups community (https://dgroups.org/fao/fisnet accessed on 13 September 2022) that provided us with a platform for communication. We thank Bruce D. Moltzan from the USDA (United States Department of Agriculture) Forest Service and Shiroma Sathyapala from the FAO Forestry Office for their help in international communication. We thank Qidi Hou from Beijing Forestry University for providing us with the literature on nematode biology.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Steiner, G.; Buhrer, E.M. Aphelenchoides xylophilus n. sp., a nematode associated with blue-stain and other fungi in timber. J. Agric. Res. 1934, 48, 949–951. [Google Scholar]
- Bowers, W.; Hudak, J.; Raske, A.; Magasi, L.; Myren, D.; Lachance, D.; Cerezke, H.; Van Sickle, G. Host and Vector Surveys for the Pinewood Nematode, Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle (Nematoda: Aphelenchoididae) in Canada; Great Lakes Forestry Centre: Sault Ste. Marie, ON, Canada, 1992. [Google Scholar]
- Dwinell, L. First report of pinewood nematode (Bursaphelenchus xylophilus) in Mexico. Plant Dis. 1993, 77, 846. [Google Scholar] [CrossRef]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Zhao, B.G.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar] [CrossRef]
- Yi, C.K.; Byun, B.H.; Park, J.D.; Yang, S.; Chang, K.H. First finding of the pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle and its insect vector in Korea. Res. Rep. For. Res. Inst. 1989, 38, 141–149. (In Korean) [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, causal agent of pine wilt disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Ruan, X.S. Occurrence and control of pine wood nematode in Taiwan. Plant Quar. 1996, 18, 86–87. (In Chinese) [Google Scholar]
- Winney, R. Preliminary observation on pine wood nematode in Hong Kong. Guangdong For. Sci. Technol. 1987, 16, 46–50. (In Chinese) [Google Scholar]
- Sun, Y.C. Pine wood nematode found in Sun Yat-sen Mausoleum, Nanjing. J. Jiangsu For. Sci. Technol. 1982, 9, 47. (In Chinese) [Google Scholar]
- Ye, J.R. Epidemic status of pine wilt disease in China and its prevention and control techniques and counter measures. Sci. Silvae Sin. 2019, 55, 1–10. (In Chinese) [Google Scholar]
- Zhang, X.; Zhao, J.J.; Yan, J.; Fang, G.F.; Huang, J.X. Economic loss assessment of pine wilt disease in Mainland China in 2017. J. Beijing For. Univ. 2020, 42, 96–106. (In Chinese) [Google Scholar]
- Zhao, J.J.; Huang, J.X.; Yan, J.; Fang, G.F. Economic loss of pine wood nematode disease in Mainland China from 1998 to 2017. Forests 2020, 11, 1042. [Google Scholar] [CrossRef]
- Song, Y.S.; Zang, X.Q. Adaptability analysis of pine wood nematode in China and preliminary study on quarantine countermeasures. For. Pest Dis. 1989, 8, 38–41. (In Chinese) [Google Scholar]
- Pan, L.; Cui, R.; Li, Y.; Feng, Y.; Zhang, X. Investigation of pinewood nematodes in Pinus tabuliformis Carr. under low-temperature conditions in Fushun, China. Forests 2020, 11, 993. [Google Scholar] [CrossRef]
- Yu, H.Y.; Wu, H. New host plant and new vector insect of pine wood nematode found in Liaoning Province. For. Pest Dis. 2018, 37, 61. (In Chinese) [Google Scholar]
- Yu, H.Y.; Wu, H.; Zhang, X.D.; Wang, L.M.; Zhang, X.F.; Song, Y.S. Preliminary study on Larix spp. infected by Bursaphelenchus xylophilus in natural environment. For. Pest Dis. 2019, 38, 7–10. (In Chinese) [Google Scholar]
- Li, M.; Li, H.; Sheng, R.C.; Sun, H.; Sun, S.H.; Chen, F.M. The First record of Monochamus saltuarius (Coleoptera; Cerambycidae) as vector of Bursaphelenchus xylophilus and its new potential hosts in China. Insects 2020, 11, 636. [Google Scholar] [CrossRef]
- Lv, Q.; Wang, W.D.; Liang, J.; Yan, D.H.; Jia, X.Z.; Zhang, X.Y. Potential suitability assessment of Bursaphelenchus xylophilus in China. For. Res. 2005, 18, 460–464. (In Chinese) [Google Scholar]
- Pan, H.W. Study on the potential geographic distribution of Bursaphelenchus xylophilus in. China. Dissertation, China Academy of Forestry, Beijing, China, 2009. (In Chinese). [Google Scholar]
- Zhang, X.Y.; Lv, Q.; Feng, Y.M.; Shi, L.; Liang, J.; Yan, D.H.; Liu, Z.Y. Risk Assessment and Countermeasures of Pine wood Nematode Disease in China; Science Press: Beijing, China, 2011. (In Chinese) [Google Scholar]
- He, S.Y.; Wen, J.B.; Luo, Y.Q.; Zong, S.X.; Zhao, Y.X.; Han, J. The predicted geographical distribution of Bursaphelenchus xylophilus in China under climate warming. Chin. J. Appl. Entomol. 2012, 49, 236–243. (In Chinese) [Google Scholar]
- Zhao, J.; Han, X.; Shi, J. Potential distribution of Bursaphelenchus xylophilus in China due to adaptation cold conditions. J. Biosaf. 2017, 26, 191–198. (In Chinese) [Google Scholar]
- Yu, Z.J.; Li, S.; Zhou, Y.T.; Zhou, R.L.; Wang, Y.X. Spatial estimation and prediction of suitable distribution of Bursaphelenchus xylophilus with different warming modes in China. J. Northeast For. Univ. 2018, 46, 85–91. (In Chinese) [Google Scholar]
- Han, Y.Y.; Wang, Y.; Xiang, Y.; Ye, J.R. Prediction of potential distribution of Bursaphelenchus xylophilus in China based on Maxent ecological niche model. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2015, 39, 6–10. (In Chinese) [Google Scholar]
- Tang, X.; Yuan, Y.; Li, X.; Zhang, J. Maximum entropy modeling to predict the impact of climate change on pine wilt disease in China. Front Plant Sci 2021, 12, 652500. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sun, H.; Zhou, Y.T.; Li, X.D.; Yu, Z.J.; Dong, Z.H. Occurrence of major forest pests in 2021 and prediction of occurrence trend in 2022. For. Pest Dis. 2022, 41, 44–47. (In Chinese) [Google Scholar]
- Song, Y.S. Pine Wilt Disease in China: Thirty Years Review and Thinking; Northeast Forestry University Press: Harbin, China, 2013. (In Chinese) [Google Scholar]
- General Station of Forest and Grassland Pest Control of National Forestry and Grassland Administration; Nanjing Forestry University. Occurrence Regularity and Control Techniques of Pine Wilt Disease in China; China Forestry Publishing House: Beijing, China, 2019. (In Chinese) [Google Scholar]
- Ding, X.; Guo, Y.; Ye, J.; Wu, X.; Lin, S.; Chen, F.; Zhu, L.; Huang, L.; Song, X.; Zhang, Y.; et al. Population differentiation and epidemic tracking of Bursaphelenchus xylophilus in China based on chromosome-level assembly and whole-genome sequencing data. Pest Manag. Sci. 2022, 78, 1213–1226. [Google Scholar] [CrossRef]
- Kanzaki, N.; Giblin-Davis, R.M. Diversity and plant pathogenicity of Bursaphelenchus and related nematodes in relation to their vector bionomics. Curr. For. Rep. 2018, 4, 85–100. [Google Scholar] [CrossRef]
- Ryss, A.; Vieira, P.; Mota, M.; Kulinich, O. A synopsis of the genus Bursaphelenchus Fuchs, 1937 (Aphelenchida: Parasitaphelenchidae) with keys to species. Nematology 2005, 7, 393–458. [Google Scholar] [CrossRef]
- Wang, H.Y. Biodiversity and Biogeography of the Genus Bursaphelenchus Fuchs, 1937 (Nematoda: Parasitaphelenchidae). Doctoral Dissertation, Fujian Agriculture and Forestry University, Fuzhou, China, 2008. (In Chinese). [Google Scholar]
- Aikawa, T. Transmission biology of Bursaphelenchus xylophilus in relation to its insect vector. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 123–138. [Google Scholar]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Zhang, X.; Wei, Y.; Zhou, J.; Zhang, W.; Qin, P.; Chinta, S.; Kong, X.; Liu, Y.; Yu, H.; et al. Ascarosides coordinate the dispersal of a plant-parasitic nematode with the metamorphosis of its vector beetle. Nat. Commun. 2016, 7, 12341. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, F.X.; Hasegawa, K.; Mota, M.; Vicente, C.S. Bacterial role in pine wilt disease development–review and future perspectives. Environ. Microbiol. Rep. 2015, 7, 51–63. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Inacio, M.L. Insights into the role of fungi in pine wilt disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.H.; Zhang, B.; Sun, J.H. Research progress on the interaction between associated microbes and pine wood nematode-vector beetle complex. For. Pest Dis. 2022, 41, 30–37. (In Chinese) [Google Scholar]
- Proenca, D.N.; Grass, G.; Morais, P.V. Understanding pine wilt disease: Roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. Microbiologyopen 2017, 6, e00415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.N.; Wang, L.; Shi, X.F.; Luo, L.; Ye, J.R. Negative effects of free-living nematodes on the populations of Bursaphelenchus xylophilus in dead pine trees. Biol. Control 2022, 168, 104858. [Google Scholar] [CrossRef]
- Akbulut, S.; Stamps, W.T. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Kwon, H.J.; Jung, J.K.; Jung, C.; Han, H.; Koh, S.H. Dispersal capacity of Monochamus saltuarius on flight mills. Entomol. Exp. Appl. 2018, 166, 420–427. [Google Scholar] [CrossRef]
- David, G.; Giffard, B.; Piou, D.; Jactel, H. Dispersal capacity of Monochamus galloprovincialis, the European vector of the pine wood nematode, on flight mills. J. Appl. Entomol. 2014, 138, 566–576. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhang, R.Z.; Chen, J.Y. Species and their dispersal ability of Monochamus as vectors to transmit Bursaphelenchus xylophilus. J. Zhejiang A F Univ. 2007, 24, 350–356. (In Chinese) [Google Scholar]
- Li, H. Identification and Pathogenicity of Bursaphelenchus Species (Nematoda: Parasitaphelenchidae). Doctoral Dissertation, Ghent University, Ghent, Belgium, 2008. [Google Scholar]
- Futai, K.; Sutherland, J.R. Pathogenicity and attraction to host extracts of Canadian pinewood nematodes: Studies with Scots pine, western larch, and black spruce seedlings. Can. J. For. Res. 1989, 19, 1256–1261. [Google Scholar] [CrossRef]
- Kiyohara, T.; Bolla, R. Pathogenic variability among populations of the pinewood nematode, Bursaphelenchus xylophilus. For. Sci. 1990, 36, 1061–1076. [Google Scholar] [CrossRef]
- Liu, K.C.; Zeng, F.L.; Ben, A.L.; Han, Z.M. Pathogenicity and repulsion for toxin-producing bacteria of dominant bacteria on the surface of American pine wood nematodes. J. Phytopathol. 2017, 165, 580–588. [Google Scholar] [CrossRef]
- Jiao, G.Y.; Shen, P.Y.; Li, H.M.; Xu, P.F. Study on pathogenicity of Japanese pine wood nematode strain and Nanjing pine wood nematode strain to Cedrus deodara and Pinus massoniana. Plant Quar. 1996, 18, 193–195. (In Chinese) [Google Scholar]
- Ju, Y.W.; Zhao, B.G.; Pu, C.H. Different virulence of pine wood nematode from Japan and China on Cedrus deodara. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2007, 31, 139–140. (In Chinese) [Google Scholar]
- Zheng, J.R. Discussion on the Mechanism of Cedar’s Immunity to Pine Wood Nematode under Natural Conditions in China. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2007. (In Chinese). [Google Scholar]
- Zhen, W.; Chun, Y.W.; Zhe, M.F.; Dong, L.Z.; Lei, L.; Mi, R.L.; Zheng, L.; Jing, J.L.; Chang, K.S. Advances in research of pathogenic mechanism of pine wilt disease. Afr. J. Microbiol. Res. 2010, 4, 437–442. [Google Scholar] [CrossRef]
- Li, Y.X.; Wang, X.; Liu, Z.K.; Zhang, X.Y. Research advance of pathogenic mechanism of pine wood nematode. For. Pest Dis. 2022, 41, 11–20. (In Chinese) [Google Scholar]
- Shinya, R.; Morisaka, H.; Takeuchi, Y.; Futai, K.; Ueda, M. Making headway in understanding pine wilt disease: What do we perceive in the postgenomic era? J. Biosci. Bioeng. 2013, 116, 1–8. [Google Scholar] [CrossRef]
- Wen, T.Y.; Wu, X.Q.; Hu, L.J.; Qiu, Y.J.; Rui, L.; Zhang, Y.; Ding, X.L.; Ye, J.R. A novel pine wood nematode effector, BxSCD1, suppresses plant immunity and interacts with an ethylene-forming enzyme in pine. Mol. Plant Pathol. 2021, 22, 1399–1412. [Google Scholar] [CrossRef]
- Wang, R.; Li, Y.; Li, D.; Zhang, W.; Wang, X.; Wen, X.; Liu, Z.; Feng, Y.; Zhang, X. Identification of the Extracellular Nuclease Influencing Soaking RNA Interference Efficiency in Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2022, 23, 12278. [Google Scholar] [CrossRef]
- Rutherford, T.; Webster, J. Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Can. J. For. Res. 1987, 17, 1050–1059. [Google Scholar] [CrossRef]
- Fan, H.J.; Guan, Z.X. Analysis on the changes of meteorological elements in Wangqing county in the last 52 years. Jilin Meteorol. 2010, 17, 15–18. (In Chinese) [Google Scholar]
- Wharton, D.A.; Goodall, G.; Marshall, C.J. Freezing survival and cryoprotective dehydration as cold tolerance mechanisms in the Antarctic nematode Panagrolaimus davidi. J. Exp. Biol. 2003, 206, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.R.; Wharton, D.A. The ability of the Antarctic nematode Panagrolaimus davidi to survive intracellular freezing is dependent upon nutritional status. J. Comp. Physiol. B 2013, 183, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Shatilovich, A.; Tchesunov, A.; Neretina, T.; Grabarnik, I.; Gubin, S.; Vishnivetskaya, T.; Onstott, T.C.; Rivkina, E. Viable nematodes from late Pleistocene permafrost of the Kolyma river lowland. Dokl. Biol. Sci. 2018, 480, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.R.; Fiedler, T.J.; Harris, T.W.; Carvalho, F.; Antoshechkin, I.; Han, M.; Sternberg, P.W.; Stein, L.D.; Chalfie, M. WormBook: The online review of Caenorhabditis elegans biology. Nucleic Acids Res. 2007, 35, D472–D475. [Google Scholar] [CrossRef]
- Szewczyk, N.J.; Mancinelli, R.L.; McLamb, W.; Reed, D.; Blumberg, B.S.; Conley, C.A. Caenorhabditis elegans survives atmospheric breakup of STS-107, space shuttle Columbia. Astrobiology 2005, 5, 690–705. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, N.; Futai, K. A PCR primer set for determination of phylogenetic relationships of Bursaphelenchus species within the xylophilus group. Nematology 2002, 4, 35–41. [Google Scholar] [CrossRef]
- Li, Z.X.; Tao, J.; Zong, S.X. Cold tolerance in pinewood nematode Bursaphelenchus xylophilus promoted multiple invasion events in mid-temperate zone of China. Forests 2022, 13, 1100. [Google Scholar] [CrossRef]
- Pan, L.; Cui, R.; Li, Y.X.; Zhang, W.; Bai, J.W.; Li, J.W.; Zhang, X.Y. Third-Stage dispersal juveniles of Bursaphelenchus xylophilus can resist low-temperature stress by entering cryptobiosis. Biology 2021, 10, 785. [Google Scholar] [CrossRef]
- Kong, Q.Q.; Ding, X.L.; Chen, Y.F.; Ye, J.R. Comparison of morphological indexes and the pathogenicity of Bursaphelenchus xylophilus in northern and southern China. Forests 2021, 12, 310. [Google Scholar] [CrossRef]
- Li, Y.X.; Zhang, X.Y. Analysis on the trend of invasion and expansion of Bursaphelenchus xylophilus. For. Pest Dis. 2018, 37, 1–4. (In Chinese) [Google Scholar]
- Yik, C.P.; Birchfield, W. Observations on the morphology of the pine wood nematode, Bursaphelenchus xylophilus. J. Nematol. 1981, 13, 376–384. [Google Scholar] [PubMed]
- Dropkin, V.; Foudin, A.; Kondo, E.; Linit, M.; Smith, M.; Robbins, K. Pinewood nematode: A threat to US forests? [Bursaphelenchus xylophilus]. Plant Dis. 1981, 65, 1022–1027. [Google Scholar] [CrossRef]
- Wingfield, M.; Blanchette, R. The pine-wood nematode, Bursaphelenchus xylophilus, in Minnesota and Wisconsin: Insect associates and transmission studies. Can. J. For. Res. 1983, 13, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Blunt, T.D.; Jacobi, W.R.; Appel, J.A.; Tisserat, N.; Todd, T.C. First report of pine wilt in Colorado, USA. Plant Health Prog. 2014, 15, 103–104. [Google Scholar] [CrossRef] [Green Version]
- Ohsawa, M.; Akiba, M. Possible altitude and temperature limits on pine wilt disease: The reproduction of vector sawyer beetles (Monochamus alternatus), survival of causal nematode (Bursaphelenchus xylophilus), and occurrence of damage caused by the disease. Eur. J. For. Res. 2014, 133, 225–233. [Google Scholar] [CrossRef]
- Haran, J.; Roques, A.; Bernard, A.; Robinet, C.; Roux, G. Altitudinal barrier to the spread of an invasive species: Could the Pyrenean chain slow the natural spread of the pinewood nematode? PLoS ONE 2015, 10, e0134126. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y. The biogeography and economic geography of the pine wood nematode in China. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2011, 35, 144–145. (In Chinese) [Google Scholar]
- Fu, L.; Li, N.; Elias, T.; Mill, R. Pinaceae. In Flora of China 4; Flora of China Editorial Committee; Science Press: Beijing, China, 1999; pp. 9–10. [Google Scholar]
- Ye, J.R.; Wu, X.Q. Research progress of pine wilt disease. For. Pest Dis. 2022, 41, 1–10. (In Chinese) [Google Scholar]
- Ge, J.J.; Bai, X.Y.; Cheng, H.R. Resistance of Cedrus deodara and Pinus taiwanensis to pine wood nematode (Bursapherenchus xylophilus). Acta Phytopathol. Sin. 1992, 23, 335–338. (In Chinese) [Google Scholar]
- Editorial Committee of FRPS Chinese Academy of Sciences. Flora Reipublicae Popularis Sinicae Volume 7: Gymnosperm; Science Press: Beijing, China, 1978. (In Chinese) [Google Scholar]
- Sutherland, J.R.; Ring, F.M.; Seed, J.E. Canadian conifers as hosts of the pinewood nematode (Bursaphelenchus xylophilus): Results of seedling inoculations. Scand. J. For. Res. 1991, 6, 209–216. [Google Scholar] [CrossRef]
- Yang, B.J.; Hu, K.J.; Wang, Q.L.; Sun, Y.L.; Wu, Z.M.; Wang, Q.M. Studies on the resistance of pine trees to pine wood nematode, Bursaphelenchus xylophilus. For. Res. 1993, 6, 249–255. (In Chinese) [Google Scholar]
- Wingfield, M.; Blanchette, R.; Kondo, E. The pine wood nematode on balsam fir in Minnesota. Phytopatholgy 1982, 72, 965. [Google Scholar]
- Kondo, E.; Foudin, A.; Linit, M.; Smith, M.; Bolla, R.; Winter, R.; Dropkin, V. Pine wilt Disease-Nematological, Entomological, and Biochemical Investigations; University of Missouri-Columbia Agricultural Experiment Station: Columbia, SC, USA, 1982. [Google Scholar]
- Kasuga, H.; Togashi, K. Nine-month persistence of Bursaphelenchus xylophilus in living Abies sachalinensis and Picea jezoensis seedlings and nematode reproduction in stem sections. Nematology 2012, 14, 547–554. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Lima, M.; Vasconcelos, M.W. Susceptibility evaluation of Picea abies and Cupressus lusitanica to the pine wood nematode (Bursaphelenchus xylophilus). Plant Pathol. 2013, 62, 1398–1406. [Google Scholar] [CrossRef]
- Editorial Committee of China Forest. China Forest Volume 1 General; China Forestry Publishing House: Beijing, China, 1997. (In Chinese) [Google Scholar]
- Editorial Committee of China Forest. China Forest Volume 2 Coniferous Forest; China Forestry Publishing House: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Cesari, M.; Marescalchi, O.; Francardi, V.; Mantovani, B. Taxonomy and phylogeny of European Monochamus species: First molecular and karyological data. J. Zool. Syst. Evol. Res. 2005, 43, 1–7. [Google Scholar] [CrossRef]
- Wang, Q. Biogeography study of Monochamus Guér. Sci. Silvae Sin. 1988, 24, 297–302. (In Chinese) [Google Scholar]
- Zheng, S.Z.; Zhang, K.; Yang, X.J.; Xu, M.; An, Y.L. Sequence comparison and phylogenetic analysis of mt DNA CO I gene sequence fragments in Monochamus species (Coleoptera: Cerambycidae). Chin. J. Appl. Entomol. 2015, 52, 370–381. (In Chinese) [Google Scholar]
- General Station of Forest and Grassland Pest Control of National Forestry and Grassland Administration. Forest pest in China 2014–2017 National Forest Pest Survey Results; China Forestry Publishing House: Beijing, China, 2019. (In Chinese) [Google Scholar]
- General Station of Forest Pest Control of State Forestry Administration. Overview of Forestry Pests in China: Compilation of National Forestry Pest Survey Results from 2003 to 2007; China Forestry Publishing House: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Department of Forestry Administration and Protection of the Ministry of Forestry of the People’s Republic of China. List of Forest Pest and Disease Survey in China; Department of Forestry Administration and Protection of the Ministry of Forestry: Beijing, China, 1988. (In Chinese)
- Shanghai Landscape Gardening Research Institute. Survey of Plant Diseases, Pests and Natural Enemies of Garden Plants in China and Study on Quarantine Pests (II); Shanghai Landscape Gardening Research Institute: Shanghai, China, 1987. (In Chinese) [Google Scholar]
- Hua, L.Z. List of Cerambycidae Insects in China; Sun Yatsen University Press: Guangzhou, China, 1982. (In Chinese) [Google Scholar]
- Chen, S.X.; Xie, Y.Z.; Deng, G.F. Economic Insects of China Vol. 1 Coleoptera Cerambycidae; Science Press: Beijing, China, 1959. (In Chinese) [Google Scholar]
- Pu, F.J. Economic Insects of China Vol. 19 Coleoptera Cerambycidae 2; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]
- Jiang, S.N.; Pu, F.J.; Hua, L.Z. Economic Insects of China Vol. 35 Coleoptera Cerambycidae 3; Science Press: Beijing China, 1985. (In Chinese) [Google Scholar]
- Wang, Z.M.; Pi, Z.Q.; Hou, B. Monochamus alternatus found in Jilin Province. For. Pest Dis. 2006, 25, 35. (In Chinese) [Google Scholar]
- Pajares, J.A.; Alvarez, G.; Hall, D.R.; Ibarra, N.; Hoch, G.; Halbig, P.; Cocos, D.; Johansson, H.; Schroeder, M. Attractants for management of the pine sawyer beetle Monochamus sutor, a potential vector of Bursaphelenchus xylophilus. J. Appl. Entomol. 2017, 141, 97–111. [Google Scholar] [CrossRef]
- Schroeder, M. Trapping strategy for Monochamus sutor and Monochamus galloprovincialis: Potential vectors of the pine wood nematode in Scandinavia. Agric. For. Entomol. 2019, 21, 372–378. [Google Scholar] [CrossRef]
- Wang, K.; Li, L.J.; Yu, F.; Wang, L.G.; Yang, H.J. Analysis and recommendations about epidemic situation of intercepted longhorn beetles at China’s ports. For. Pest Dis. 2018, 37, 22–26. (In Chinese) [Google Scholar]
Figure 1.
Pine wilt disease epidemic provinces in China (2022). The orange and red-filled areas are the provinces with PWD. In the pink-filled area, PWD has been eradicated. The yellow squares are marked as the most frontier epidemic areas in Northwest and Northeast China.
Figure 1.
Pine wilt disease epidemic provinces in China (2022). The orange and red-filled areas are the provinces with PWD. In the pink-filled area, PWD has been eradicated. The yellow squares are marked as the most frontier epidemic areas in Northwest and Northeast China.

Figure 2.
Relationship of PWNs and the genera of Pinaceae in China.


{kind=link}
{kind=link}
Table 1.
Reports on the isolation of PWNs from Picea and Abies trees in the field and the death of Picea and Abies by artificial inoculation with PWNs.
Table 1.
Reports on the isolation of PWNs from Picea and Abies trees in the field and the death of Picea and Abies by artificial inoculation with PWNs.
| Country | Isolation | Seedling Inoculation |
|---|---|---|
| USA (Origin) | Picea glauca, Picea pungens, Abies balsamea [33,84,85] | |
| Canada (Origin) | Picea glauca, Picea mariana, Picea rubens, Abies balsamea [2] | Picea engelmannii, Picea glauca, Picea mariana, Picea rubens, Picea sitchensis, Abies amabilis, Abies balsamea, Abies grandis [82,83] |
| Japan (Invasive) | Picea abies, Abies firma [4,5] | Picea abies, Picea jezoensis, Picea mariana, Abies homolepis, Abies sachalinensis [5,86] |
Table 2.
Conifer tree species with the lowest distribution at an altitude below 1000 m in Northeast and Northwest China.
Table 2.
Conifer tree species with the lowest distribution at an altitude below 1000 m in Northeast and Northwest China.
| Species | Distribution in Northeast (NE) and Northwest (NW) China | Distribution | Lowest Distribution Altitude (Meters) |
|---|---|---|---|
| Pinus densiflora var. ussuriensis | NE | Heilongjiang Province | 50 |
| Pinus koraiensis | NE | Lesser Khingan (Heilongjiang Province), Wanda Mountains (Heilongjiang Province), Zhangguangcai Range (Heilongjiang and Jilin provinces), Changbai Mountain (Jilin Province) | 50 |
| Pinus sylvestris var. sylvestriformis | NE | Changbai Mountain | 630 |
| Pinus thunbergii | NE | Liaoning Province | 400 |
| Larix gmelinii | NE | Greater Khingan (Heilongjiang Province), Lesser Khingan | 300 |
| Larix olgensis | NE | Changbai Mountain | 500 |
| Picea jezoensis | NE | Greater Khingan, Lesser Khingan, Zhangguangcai Range, Changbai Mountain | 700 |
| Picea koraiensis | NE | Greater Khingan, Lesser Khingan, Wanda Mountains, Zhangguangcai Range, Changbai Mountain | 300 |
| Abies nephrolepis | NE | Lesser Khingan, Zhangguangcai Range, Changbai Mountain | 500 |
Table 3.
Monochamus species feeding on conifers in China.
| Species | Distribution in Northeast (NE) and Northwest (NW) China | Relationship with PWN (Pine Wood Nematode) |
|---|---|---|
| Monochamus alternatus | NE and NW | Confirmed vector in China [43] |
| Monochamus basifossulatus | No distribution | |
| Monochamus galloprovincialis | NE | Confirmed vector in Europe [43] |
| Monochamus guerryi | No distribution | |
| Monochamus saltuarius | NE and NW | Confirmed vector in China [19] |
| Monochamus sartor urussovii | NE and NW | |
| Monochamus sutor | NE and NW |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, Q.; Zhang, X.; Li, J.; Ren, J.; Ren, L.; Luo, Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests 2023, 14, 174. https://doi.org/10.3390/f14020174
AMA Style
Xu Q, Zhang X, Li J, Ren J, Ren L, Luo Y. Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review. Forests. 2023; 14(2):174. https://doi.org/10.3390/f14020174
Chicago/Turabian StyleXu, Qinwang, Xuejiao Zhang, Jiaxing Li, Jiaru Ren, Lili Ren, and Youqing Luo. 2023. "Pine Wilt Disease in Northeast and Northwest China: A Comprehensive Risk Review" Forests 14, no. 2: 174. https://doi.org/10.3390/f14020174
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.