
Shoot Phenology as a Driver or Modulator of Stem Diameter Growth and Wood Properties, with Special Reference to Pinus radiata

Scion (New Zealand Forest Research Institute Ltd.), Private Bag 3020, Rotorua 3046, New Zealand
Forests 2023, 14(3), 570; https://doi.org/10.3390/f14030570
Submission received: 12 December 2022
/
Revised: 23 February 2023
/
Accepted: 2 March 2023
/
Published: 13 March 2023
(This article belongs to the Special Issue Intrinsic Regulation of Diameter Growth in Woody Plants)
Abstract
:Seasonal phenology is expressed in the annual rhythms of growth and quiescence, which may range from being sharply defined to weakly quantitative. These rhythms, both vegetative and reproductive, are a key aspect of a plant’s survival strategy. They enable the plant to both survive seasonal stresses and take advantage of favourable conditions, while making pollination efficient through synchronised flowering. Maturation in woody perennials causes some ontogenetic modulation of seasonal phenology. Shoot phenology is driven by various environmental cues, notably temperatures and daylength, with shoot extremities often being prime receptors. The phenology of shoot extremities is in turn seen as a strong driver of cambial activity, which itself is an aspect of shoot phenology and the basis of stem diameter growth. The aspects of cambial activity reflecting hoot–tip phenology primarily involve the timing of xylem formation and the anatomical and physical properties of xylem cells. The actual amount of diameter growth, however, is governed much more by other factors, notably the growth potential of the species, the space for the tree to grow in, and how favourable local conditions are for growth. Somehow, all tree species allocate resources to shoot extension and diameter growth to represent viable adaptive strategies, although there is no identified role of phenology per se in such allocation. Among species, Pinus radiata is extremely important commercially, largely through a high growth potential that reflects distinctive phenology. Much research on the species provides a basis for linking its phenology and shoot–tip architecture to seasonal timing of diameter growth and fine details of within-ring variation in xylem cell properties. This provides a baseline for comparison with many other tree species, conifers and hardwoods. Selected examples are considered to illustrate the range of adaptive strategies. Regulation of diameter growth and internal variation in wood properties reflects the response to competitive pressures. Modifying the regulation to optimise wood production and quality poses a great challenge for future domestication.
1. Introduction
In growing forest trees for their wood, we always want to maximise both the quantity and quality of the wood, while minimising the risks from biotic and abiotic agents. Optimisation, however, is typically complicated by some trade-offs between the quality and quantity of wood, [1] and the fact that the criteria of quality can often be specific to end uses. Additionally, despite some scope for fine-tuning wood properties for end products and their uses, there will be fundamental limitations that constrain the processing and end uses of the wood of individual species.
Trees, as long-lived woody perennials, must cope with whatever seasonal fluctuations there are. In temperate to boreal zones, seasons are dominated by temperatures and daylengths. In the tropics and Mediterranean to subtropical zones, seasons are typically defined much more by patterns of precipitation. Moreover, whatever the latitude, increasing elevation above sea level will lower temperatures, accentuating winter minima and mitigating summer maxima and typically increasing precipitation. These seasonal fluctuations, which can often pose additional adaptive challenges for plants, have various influences on the diameter growth entailed in wood formation. These influences govern the patterns of variation in anatomical and physical properties within annual growth rings, whereas the amount of wood produced is governed much more by the space and other resources available to the tree. The intra-ring variation in detailed properties, however, can be of major technical importance.
Seasonal phenology, and the various features that it involves, is surely crucial to plants remaining viable and competitive in their various environments, as reviewed by Park and Post [2]. The manifestations are many and varied. Those in spring are often the clearest and as such may be the starting point for characterisation, even though they are only part of a repeating annual cycle. Moreover, phenology involves not only vegetative processes but also reproduction, in flowering through to ripening and dispersal of seed, a matter that I will address separately.
General aspects of phenology are reviewed. Then Pinus radiata is considered, mainly in terms of its importance, its phenology, the environmental cues and other factors, environmental influences on phenotype, and how the phenology governs the within-ring profile in wood properties. To place P. radiata in perspective, within-ring profiles of a selection of other species are reviewed. As an overarching issue for tree diameter growth, meeting both competitive needs and mechanical stability is considered with implications for intensive domestication. Finally, some conclusions are drawn. Full, balanced literature coverage is not attempted.
2. General Aspects of Phenology
2.1. Processes Involved
Seasonal phenology involves the timing of a wide range of processes. They typically involve alternation between periods of quiescence and active growth. The vegetative processes include bud break and the subsequent elongation of shoots and leaves, through to the winter bud set, with concomitant changes in tolerances of factors such as cold and drought. In trees that are deciduous, the complete leaf fall that this entails represents a major phenological event. Not directly visible are the processes involving cambial activity, from the onset of xylem cell formation through the change from producing earlywood (EW) to latewood (LW) with the cessation at the end of the season. The most common feature differentiating LW from EW is higher density. In softwoods, the LW tends to define the annual ring visually, whereas in ring-porous hardwoods the earlywood does so. The higher density of LW in softwoods variously reflects the reduced radial dimensions of tracheids, e.g., [3], and/or thicker walls, e.g., [4]. In ring-porous hardwoods, vessels predominate in the EW, with fibres tending to predominate in the LW. However, EW often intergrades into LW. The radial density profiles within annual rings are now tracked radiometrically, generally using X-rays, e.g., [5,6,7].
Less studied is the seasonal course of producing phloem elements from the cambium. Also less studied, and even more cryptic, are the seasonal phenomena of root growth; however, it seems safe to assume that they largely reflect the above-ground phenology, which copes with much greater seasonal fluctuations. The strength and nature of seasonal variations vary widely among different parts of the world. Also, the architecture of shoot extension varies widely among taxa and environments. So too, do the strength and nature of plant seasonal phenology.
Reproductive processes can often be the most conspicuous manifestations of phenology in flowering (“Flowering” is used in a broad sense of the term because gymnosperms are not considered to be flowering plants in the sense that angiosperms are) and/or fruit ripening of some plants. The timing of flowering is doubtless governed in evolutionary adaptation by a number of factors, including the risk of frost or other seasonal damage, and the need to complete seed development and dispersal. Furthermore, close synchronicity of flowering is clearly of special importance for efficient pollination. Depending on the physical architecture of shoots and flowering structures, the degree of coupling between vegetative and reproductive can vary widely among groups of plant species. For instance, with pines (Pinus spp.), vegetative and reproductive phenology are quite closely coupled through developmental architecture as well as in seasonal timing [8]. By contrast, in tropical hardwoods that show cauliflory (flowering low on the trunk rather than on shoot tips) such coupling seems minimal or absent.
2.2. The Biotic Overlay
Plant phenology, while representing adaptation to seasonal fluctuations, does not always do so in an intuitively straightforward way. Biotic factors, which often have their own seasonal influences, are thought to contribute [9]. Specifically, the presence of plant pathogens has been postulated to contribute to the evolutionary persistence and/or seasonal timing of deciduousness, which would be an avoidance mechanism. Such a mechanism may operate in both tropical and temperate zones. Against herbivory, close synchronisation and a brief period of shoot flushing are postulated to confer protection by “predator satiation” (or “predator swamping”). This is more pronounced in tropical or subtropical climates [10,11], where seasons are defined more by rainfall than by temperature and there is no strong confounding with seasonal responses to temperatures.
2.3. Environmental Cues
Expression of seasonal phenology is governed by a range of environmental cues. In temperate climates, temperature generally figures prominently among the cues. In spring, the most recognisable cue for bud burst is usually rising temperature [12], although its efficacy may depend on a chilling (vernalisation) and/or photoperiodic (daylength) requirement having been met. Daylength is often a strong regulator of shoot elongation, e.g., [12,13,14], with the paradoxical feature that it can be longest at the high latitudes that limit potential growth. For setting buds and frost hardening in autumn, shortening days and/or lowering temperatures can variously be the cues. Less defined and less consistent is a role of drought in modifying and sometimes establishing seasonal phenology. In Mediterranean climates there is the complication that, in respect of adaptation, seasons are primarily defined by both temperature and precipitation levels. However, the cues provided by one of these factors may govern adaptation to coping with the other, not to the exclusion of photoperiod as a cue.
In tropical regions, where seasons are defined mainly by precipitation, a presumption is that the resulting water potential is typically involved in somehow triggering changes in phenological state. However, soil moisture status almost anywhere will be associated with variations in other features of the soil environment. For instance, drying of the upper soil layers, where key nutrients tend to be concentrated [15], can affect nutrient availability. In tropical regions, any photoperiodic cues will be subtle and potentially very complex. In equatorial regions, where seasonal fluctuations in precipitation are minor, identification of cues for seasonal phenology can be particularly challenging. For instance, Borchert et al. [16] proposed a photoperiodic model to explain the phenology of a group of peri-equatorial plants, but later (Calle et al. [17] and Borchert et al. [18]) proposed an alternative model based on insolation levels.
There are other complications in the adaptive bases for seasonal phenology in the tropics. Some of the phenology can be very counterintuitive, as with trees that produce fresh foliage in the dry season, suggesting a defence against fungal/oomycete pathogens [9]. In other cases, production of fresh shoots and foliage occurs during very brief periods, suggesting a defensive strategy of “predator satiation” (or “predator swamping”) against herbivory.
2.4. Ontogenetic Aspect
Plant phenology is generally deemed, at least tacitly, to be essentially seasonal. Yet there is typically an overlay of ontogenetic phenology involving maturation that has often been termed either phase change, e.g., [19,20], or physiological ageing. After a plant germinates from seed it undergoes various changes in anatomy and morphology. Most specific, perhaps, is the acquisition of reproductive competence, prior to which none of the standard environmental cues will induce flowering. Vegetatively, the maturation state can involve large differences in shoot morphology between juvenile and adult phases, a phenomenon termed heteroblasty. Extreme heteroblasty in some New Zealand woody perennials has been postulated as a defence against browsing by large flightless birds [21,22]. Of more relevance, however, is that ontogenetic phenology can be related to seasonal phenology, with the adult growth phase being associated with stricter seasonal phenology, e.g., [8,23].
2.5. Underlying Mechanisms
The growth of woody perennials almost always arises in shoot extension, for which the environmental cues may be known and reasonably understood. Yet most of the growth that occurs, at least in accumulating biomass, is in the cambium and its immediate derivatives. Cambial behaviour, however, typically reflects the behaviour of the shoot extremities that appear to be the main receptors of environmental cues [13,19], subject to the availability of photosynthate. The classical model, which I take as the starting point, is that of P.R. Larson [24,25,26]. Developed from a comparatively simple system, namely that of extension and radial growth of Pinus resinosa Sol. ex Aiton, Larson associated EW formation with the period of active extension growth, with LW production following. This was readily explained, even then, in terms of active shoot extension being especially conducive to auxin production, the basipetal and geotropic translocation of auxin, and the demonstrated influence of auxin on formation and characteristics of xylem cells [27,28]. Quantitative demonstration of such an effect, however, was less straightforward. Reliable assays are made difficult by steep, very fine scale gradients in auxin concentrations. Moreover, the fluxes and/or turnover of phytohormones can be more influential than assayable concentrations. Although other phytohormones are now recognised as being needed for normal xylogenesis (review by Savidge [28]), the pre-eminent role of auxin in linking shoot extension to EW–LW differentiation remains essentially unchallenged.
2.6. Differentiating between Earlywood (EW) and Latewood (LW)
Whereas the period of active shoot extension, or flushing, is often confined, the typical density profile for a softwood represents a more or less smooth radial progression [5]. Beginning low, the density often increases progressively more steeply to a maximum towards the outer margin of the ring. That means that demarcation between EW and LW is always in some degree arbitrary, as noted by Harris [29]. So too, in consequence, are the calculations of EW: LW ratio and the ratio of EW and LW densities. Indeed, the contrast between maximum and minimum within-ring densities may be more informative. Also, the curvilinear progression points to the decline in the phytochemical signal for EW production continuing after shoot and foliage elongation is complete. As addressed later, the magnitude of EW–LW differentiation is often much less (sometimes negligible) in hardwoods and the nature of differentiation is often very different [5].
2.7. The Roles of Xylem
The prime roles of the xylem, which constitutes the wood, are mechanical and hydraulic, e.g., [30]. The latter is dominated by translocation of water and dissolved nutrients from the roots to the crown. Key features of the hydraulic system are conductivity, resistance to air embolism that can block translocation (or “safety margin”) and capacitance. There tends to be a trade-off between the first two, conductivity being favoured by wide tracheids or vessels and resistance to embolism by narrow translocatory cells. In this connection, within-ring variations are of interest. Capacitance relates to the ability of the xylem to buffer the crown against short-term (often diurnal) fluctuations in water potential, being favoured by depth of sapwood cells and moderate conductivity. Safety margins tend to be especially important in very tall trees.
3. Pinus radiata—A Classical Case in Point
3.1. Introduction
Pinus radiata D.Don, or radiata pine or Monterey pine (Figure 1 and Figure 2), has gone from being an obscure Californian conifer to becoming a major commercial timber species [31,32,33,34,35].
The species has a very restricted natural range. The full natural extent was barely 10,000 ha, which has been reduced to about 5000 ha by human activity. There are five discrete natural populations (Table 1). Three are on the Californian mainland, at latitudes 35½–37 °N, and two are on Mexican islands Guadalupe at 29 °N and Cedros at 28 °N. All populations are located in a special, highly localised variant of the Mediterranean climate. It is characterised by cool sea fogs in summer coming off a cold ocean current to produce remarkably copious fog drip from tree crowns. The fog drip compensates for an almost complete lack of summer rainfall. Nowhere does the species occur naturally more than about 5 km from the sea. Although the native climates are almost never very closely matched elsewhere, the species thrives over large areas in other parts of the world, mostly in the Southern Hemisphere with mild oceanic climates. It is now grown over an area of some 4 million ha, supporting major industries on a scale far out of proportion to that area. The two countries growing it on the largest scale are Chile and New Zealand, followed by Australia and then Spain. Indeed, it is generally the utility softwood of choice for planting wherever it thrives. Domesticated stocks of the species have evidently descended almost entirely from the two northernmost natural populations, Monterey (Lat. 36½ °N and Año Nuevo (Lat. 37 °N), with a disproportionate ancestral contribution from the much smaller Año Nuevo population [34,36]. These domesticated stocks have been the subject of almost all the research on the species.
The reasons for the species’ popularity include very rapid growth, ease of raising in the nursery and transplanting, and the modest soil requirements and amenable wood properties of a typical pine (Pinus). Very helpful is the abundant genetic variability that has allowed successful genetic improvement programmes [34]. Mainly because of its commercial importance, much research has been conducted on many aspects of its biology, effectively making it an important “model species” among woody perennials. The rapid growth stems largely from a very long, opportunistic growing season [32,34,37].
3.2. Primary Environmental Determinants of Diameter Growth
In many tree species, the rate of diameter growth is constrained by growth potential, which can be limited by seasonal activity that reflects phenology. Trade-offs with wood density may also influence diameter growth. In P. radiata, the opportunistic growing season means that its phenology does not appear to be per se a primary regulator of longer-term diameter growth. Such growth is subject to the general influences of temperature, soil moisture and nutrients, and insolation. For the individual tree, as in most tree species, diameter growth is greatly influenced by general health and the effective growing space allowed by the level of inter-tree competition for limiting resources. Interestingly, the height and diameter growth of P. radiata show different profiles of response to temperature and general soil fertility [1,34,37]. Diameter growth and crop basal area increment tend to be affected more than height growth by N deficiency but less by low prevailing temperatures, in some trade-off with wood density. Evidently, diameter growth can be depressed by supra-optimal temperatures.
3.3. The General Phenology
The highly opportunistic growth reflects an apparent lack of any true low-temperature dormancy or of any actual chilling requirement for active growth and the pollination season to proceed [8]. This pints to a virtually total lack of what is termed endodormancy [38,39]. Somewhat unusually, among temperate-zone Pinus species and Pinaceae in general, the species often shows an intermediate phenological state. For a pine, and many other conifers, I define phenological state in terms of the balance between primordial initiation and shoot elongation, deeming a positive state to represent when the latter is dominant [8]. With many temperate-zone pines there is a sharp alternation between the two processes, elongation occurring almost entirely at the expense of primordial initiation during a brief season (positive state), followed by a period of primordial initiation that is reflected in some enlargement of a sealed bud that remains sealed over winter (all in negative state). This means a strong seasonal alternation between completely positive and completely negative states. In P. radiata, however, the phenological state shows more subtle and complex fluctuations, which I see as being particularly informative concerning influences on xylem cells.
Intensive studies of the seasonal growth of P. radiata include reports in the following [40,41,42,43,44,45,46] Specifically, the seasonal growth is characterised primarily by the following features, moisture levels permitting:
- Some growth occurring whenever prevailing temperatures exceed around 4–5 °C;
- Extension growth being more rapid, at least in relation to prevailing temperatures, during the spring months;
- Diameter growth, temperatures permitting, showing an upsurge from around the shortest day even though prevailing temperatures may still be falling.
Annual growth stages in P. radiata actually show a discordance between shoot extension and diameter growth [8,47]. For diameter growth, stages are clearly defined by the LW–EW boundary between consecutive rings and either the winter minimum or winter cessation. For shoot extension growth, however, a stage is typically reflected in a zone of compressed parastichies of bare cataphylls that subtend a series of either pollen strobili or fascicles [48]. This zone arises after the spring growth flush is competed. The picture is typically clear on laterals of low vigour. On the leader, where height growth occurs, the picture is less clear. The zone of compressed parastichies, which is usually evident [49], is sometimes absent. The latter case means no clear demarcation of annual growth stages. Above the zone, considerable shoot elongation can occur between the summer and winter solstices [8,47], this being interpreted as a “head start” on next season’s shoot elongation. (Cremer [44] termed this a “basal flush”). By comparison, pines (and other members of the Pinaceae) from cool temperate climates generally have a single cycle of shoot extension, meaning that the phenological state is positive only while the flush of elongation lasts. Outside this period, the only elongation that occurs is enlargement of the sealed terminal bud, which is accumulating and developing new primordia, meaning that there is no material basis for a corresponding discordance.
Reproductive activity is distinctively characterised by a very early but well-defined pollination season, the timing of which appears to be governed by when permissive temperatures are reached after the shortest day [8,47]. Most interestingly, female strobilus buds occurring among branch buds that elongate considerably during the autumn do not elongate until the pollination season is imminent.
In tropical latitudes, at altitudes that are cool enough for the species to tolerate, the phenology tends to become chaotic [50,51]. Most specifically, there is generally no well-defined pollination season and seed production is minimal. Vegetative growth is often characterised by prevalent “foxtailing” (or “terminal hypertrophy”), with very long lengths of unbranched leaders that are very prone to becoming damaged. However, dry seasons, if they are not too stressing for the species to thrive, can impose seasonality of growth [50]. Wood specimens from an equatorial site in Kenya, shown to me in 1962 by L. Chalk, had no clear annual rings, there being no clear EW–LW differentiation.
With the cambial activity that drives diameter growth, cool winters can cause a cessation unless prevailing temperatures are high enough to be permissive. In the cooler climates where the species is grown, this cessation can last for months. In milder climates, diameter growth can continue throughout the winter, which argues strongly against endodormancy. The upsurge that can occur in such climates from almost immediately after the shortest day [8,47] has been associated, albeit anecdotally, with a freeing of the cambium (A.G.D. Whyte pers. comm. 1965). However, there is almost invariably a sharp boundary between the LW of one ring and the EW of the next. Also, even in situations where diameter growth never quite ceases, the production of fresh xylem cells can cease for a period of time from before the shortest day [52,53,54].
These findings raise questions about the seasonal course of phloem cell formation and the nature of any coupling of it with xylem cell formation. Another unanswered question is whether cessation of xylem cell formation in late autumn still occurs before the shortest day in very mild subtropical climates. An important feature is that, within a radial file of tracheids, there is an abrupt change from the last LW cell with thick walls and modest radial width, to the first EW cell with thinner walls and greater radial width. Some staggering between adjoining radial files means that the LW–EW boundary is irregular at the individual tracheid level. How such an abrupt change can arise from an apparent continuum of cambial and immature xylem cells is another question.
In addition, there is empirically a strong positive association (R2~0.88) in P. radiata in New Zealand between wood density and mean annual temperature (Figure 14 in [53,55,56]). It is postulated that in cooler climates, where diameter growth commences later in the season, there is less time between the completion of EW formation and the end of the season available for LW formation. That, however, appears not to be the whole story because an association with temperature has been reported for both EW and LW densities (Figure 14 in [53,55,56]). However, quantifying relationships with EW and LW proportions and densities in growth rings is complicated by a combination of EW transitioning into LW and a consequent arbitrariness in defining LW [57]. Apart from wood density and its component traits, namely cross-sectional dimensions and cell wall thickness of tracheids, some other properties show strong effects of environment. For instance, tracheid length shows a similar trend [5,55] but it is weaker, which suggests some additional environmental influences. Genetic variation for wood and fibre properties is generally superimposed straightforwardly on such environmental effects [34].
3.4. Modulators of the Species’ Phenology
Before considering the phenology of P. radiata in further detail, it is appropriate to consider factors that modulate the character of its seasonal phenology. Three prominent ones are ontogeny or maturation state, soil fertility (most specifically nitrogen status), and shoot vigour as governed by position in the crown.
With the ontogenetic progression that represents maturation (in which onset of flowering is a distinctive manifestation), tree species generally begin after germination in a neutral phenological state, with shoot elongation essentially matching primordial initiation. In the Pinaceae, this is manifested by green apical tufts of primary foliage (Figure 3).
As the tree becomes older and larger, brown, scarious cataphylls eventually come to be produced in place of the green primary foliage. Long before this process is completed, however, fascicles (or bundles) of secondary foliage start to get produced in the axils of primary leaves or cataphylls. Eventually, a transition from producing green apical tufts to brown sealed buds is completed (Figure 4 and Figure 5).
In P. radiata, maturation progresses more gradually than in most pines [32,34], although the rate of progression varies both among populations and among individual genotypes [58,59,60]. As maturation proceeds, the seasonal alternations in phenological state become more pronounced, with shoot elongation peaking earlier in the season [8,23]. Onset of sexual reproduction is a late ontogenetic milestone. With the increased seasonality and earlier seasonal peaking of shoot elongation, some modification of the within-ring profile of fibre properties can be expected. More important though, seems to be the influence of maturation state on fibre properties [61]. The increasing level of maturation is clearly reflected in how wood properties, for a given ring number from the pith, change with increasing height up the bole. With maturation, fibre coarseness and length increase, as do grain spirality in corewood (so-called juvenile wood) and stiffness (modulus of elasticity). Also, the propensity to form compression wood decreases [62]. In loblolly pine (P. taeda L.) the pattern is broadly similar [61].
The phenological state in P. radiata shoots can also vary within the crown of an individual tree [8]. In young trees, such variation can arise incidental to within-crown variation in maturation state, with the leader and upper branches being less juvenile than the tips of lower branches. This reflects how the maturation state is governed mainly by distance along shoot axis from the original root collar [61]. Of greater perceived importance, however, is variation in phenological state according to position in crown and vigour of shoots. The leader and upper laterals tend to have longer extension seasons than do less vigorous shoots. They thus spend less time in a negative phenological state and more in a positive or roughly neutral state. This has profound implications for interpreting the radial profile of fibre properties within the annual ring according to position on the bole. This will be considered in detail later. A complication for interpretation arises in the fact that, on minor branches, the initiation and development of pollen strobili at the base of the autumn bud structure can mean the greatest shoot elongation during the autumn months [42]. It is presumed, though, that the period of elongation of the fascicles and the distal part of the annual shoot bearing them is what dominates the phenological signal to the shoot cambium. Unclear, though, is the contribution to such a signal of pollen strobilus elongation before dehiscence.
The effect of soil fertility, specifically N status, on phenological state is to dampen down seasonal fluctuations in it. This is evident irrespective of maturation state. In very young seedlings, with green apical tufts instead of sealed buds, N deficiency tends to cause episodic growth, with primordial initiation alternating in time with shoot elongation [63]. With abundant N, such seedlings would grow steadily, maintaining a neutral phenological state with pimoridal initiation and shoot elongation in balance. In the mature phase, characterised by sealed apical buds, the alternation between a positive phenological state after the shortest day and a negative one after the longest day is more pronounced with low fertility. Somewhat anecdotally, Burdon [8] has observed in older trees an absence of autumn shoot elongation in the field on N-deficient sites. High soil fertility, however, appears also to have more direct effects on wood properties, increasing the incidence of compression wood and decreasing stiffness [61,64,65]. Given that compression wood tends to have reduced stiffness [66], these two effects are likely to be interdependent. The degree of interdependence, however, remains unclear.
3.5. Key Environmental Cues
Temperature appears to play an inherently simple role in the species’ phenology, in that there is no evident chilling (or vernalisation) requirement for breaking dormancy. Indeed, there is no clear indication of true dormancy in the species, in that whereas measurable growth ceases when prevailing temperatures are below 4–5 °C [40,44,54], it can resume very quickly when temperatures rise again. Exactly what the strength of a temperature cue is in terms of an integral over time is unclear, but quite crude measures such as “degree days” can be an acceptable guide. However, cool temperatures do play a role in the cold hardening of tissues [67], although photoperiod has also been implicated [68,69]. Anyway, low temperatures, by halting growth, can effectively mask expression of the underlying phenological state. Indeed, the evidence (earlier) suggests that they do not actually govern the underlying phenological state. Notably, even without cold hardening, actively growing seedlings of P. radiata can stand temperatures as low as −6 °C without visible injury. The same temperatures can severely damage at least one conifer in its flushing season (Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco)) with much greater winter hardiness [70].
The photoperiod undoubtedly plays an important role in governing phenological state0 [8,47]. The species appears to be a variant of the long-day plant category. Although some laboratory studies have pointed to a classical long-day plant model [8,71], some other early experiments and the field behaviour suggested responding to daylength shift rather than to absolute daylength [8,47]. This is especially true in how an upsurge in diameter growth can occur in the field almost immediately after the shortest day and in how the timing of pollination season evidently reflects prevailing temperatures after the shortest day [8,72].
Even with experimental results that suggested an absolute daylength response, there is the possibility that absolute daylength treatments, as simulated in the climate laboratory, could have unintendedly elicited what in nature is a daylength-shift response. To account for observed field behaviour two alternative mechanisms have been proposed and discussed by Burdon [8].
- 1.
- Relatively direct perception of daylength shifts, possibly in terms of detecting departures from 24 h in dawn-to-dawn or dusk-to-dusk intervals. Such a mechanism would likely depend on both a very precisely temperature-compensated circadian clock and on the cue being cumulative over a period of time. The cumulative nature of such a response would automatically correct for weather-related fluctuations in the apparent dawn-to-dawn or dusk-to-dusk intervals. Cumulative responses to photoperiodic stimuli Figure 13 in [14] are a feature of long-day plants.
- 2.
- An absolute-daylength response, operating in conjunction with facultative progression of phenological state as primordia accumulate and develop. Thus, from the shortest day onwards the latter process is reinforced by increasing daylength stimulus. Conversely, after the longest day the facultative process continues to be offset by the effects of progressively shortening days.
The two mechanisms may not be altogether mutually exclusive. However, identifying what mechanism (or mechanisms) operate and tracing the gene expression pathways will pose a major research challenge [8].
3.6. The Radial Profile
Pinus radiata shows a strong radial progression from corewood to outerwood (so-called juvenile wood and mature wood) [58]. Density initially progresses rapidly out from the pith, approaching a roughly asymptotic value from around the 15th ring, placing the species in Type 1 of Schimleck et al. [73]. The progression involves various other wood properties, including tracheid length and wall thickness, grain spirality, and microfibril angle [58]. In considerable measure, this reflects an increase in the proportion of LW. However, there is also some truly ontogenetic progression up the stem, mainly in the first 3 m from ground level.
3.7. The Annual Ring Profile
Differentiating between EW and LW within the annual ring is not straightforward because the density profile typically shows a more or less smooth curve. One criterion used for differentiating them has been a specific density threshold, for example 400 or 500 kg m−3, below which is EW and above which is LW. It is convenient to apply and allows a ready cross-reference between studies using the same threshold. However, it is very arbitrary and, as Harris [29] for example noted, it does not recognise the range and pattern of ring-width variation, which I see as needed for my present purposes. Even so, the potential and actual diversity of within-ring profiles poses challenges for achieving an appropriate demarcation. Obvious alternatives are mid-range or the value at the radial mid-point within the ring, however gradual the progression might or might not be. However, our concern is with how the density profile relates to known shoot elongation phenology.
Adopting the Larson model (earlier), the profile of fibre properties across the annual growth ring reflects the phenological state of the crown distal to the cross-section through the season of active xylem growth. That state must be a summation of the states of individual shoot tips. The behaviour of P. radiata can be particularly informative because of the variation in phenological state among shoot tips and in the summation of state. The crown distal to the corewood profile contains a high proportion of shoots that are vigorous and therefore in a positive phenological state for much of the year, which is conducive to a high proportion of EW. However, any outerwood, especially in a closed stand with limited crown depth, subtends a crown that contains a much larger proportion of low-vigour branches that are only briefly in a positive phenological state.
Considering observed within-ring density profiles (Figure 6, Figure 7 and Figure 8) the following features are notable:
- 1.
- In the corewood rings, an initial zone of low density occupies around half the radius, compared with much less in the outerwood rings. This fits well with how a much higher proportion of the crown distal to the section is in an active phenological state, in accordance with the Larson model.
- 2.
- In the outerwood, the initial density within rings is higher than in the corewood. This suggests, in terms of the Larson model, that even early in the season the phytohormone signal has been attenuated by greater distance from the effective crown centre. Alternatively, there may be some more deeply epigenetic effect(s) operating.
- 3.
- In both corewood and outerwood, there is an upwards progression of density from the earlywood value towards the maximum. This accords with how the overall phenological state of the crown may be expected to become increasingly negative after the spring maximum.
- 4.
- With these trends, the curves show fluctuations that may represent some combination of biological and instrumental “noise” variation. This may account for the lack of a sharp drop at the outer margins of rings in the early traces of Harris [29].
Basically, the picture fits the Larson model, especially with the quantitative variation in phenological state that is characteristic of the species, although some additional features are suggested.
Density is far from the only property involved in the EW–LW progression. Also involved are reducing tracheid radial width and increasing wall thicknesses [4,74], which are both components of density, increasing tracheid length and increasing visibility of mild compression wood in LW. There will also be some microstructural and chemical progressions. Nonetheless, density is the most consistent and easily measured property expressing the role of phenology in the EW–LW progression.
It Is a feature of P. radiata that, even in temperate regions, the within-ring density variation is unusually modest for a conifer [4,5], which is reflected in the ease of peeling and machining. That is fully in keeping with the generally limited seasonal variation in phenological state—except with nitrogen deficiency.
3.8. Primary Environmental Determinants of Diameter Growth
The opportunistic growing season of P. radiata means that its phenology does not appear to be per se a primary regulator of longer-term diameter growth. Such regulators include, in addition to temperature, soil moisture and nutrients, insolation, and for the individual tree, general health and the growing space allowed by the level of inter-tree competition. Interestingly, height and diameter growth show different profiles of response to temperature and general soil fertility [1,32,34]. Diameter growth and carrying capacity tend to be affected more than height growth by N deficiency but less by low prevailing temperatures, albeit with some trade-offs involving wood density. Evidently, diameter growth can be depressed by supra-optimal temperatures and height growth by supra-optimal soil fertility.
3.9. Influences of Drought
Drought can obviously affect diameter growth severely [75]. Also, P. radiata is less drought tolerant than some other true pines, as is evidenced by its localised natural habitat and its failure to thrive in Mediterranean climates to which several other pines are well adapted. The phenological significance of drought per se is not totally clear, despite Lanner [51] noting that drought can impose some seasonality of behaviour in the absence of other cues. Also, drought can have indirect influences. One such is a postulated reduction of nitrogen availability. The upper soil layers, which contain a concentration of mycorrhizal roots and are most subject to moisture depletion, contain a high concentration of the available nutrients [15]. As mentioned earlier, nutrient shortage has been shown to influence phenology by accentuating phenological states.
Moderate moisture deficits need not disrupt xylem formation, even though there may be stem shrinkage involving loss of turgor in immature xylem cells during the day. That is because overnight cessation of transpiration can allow the tree to make good water deficits that arise from high daytime transpiration [76,77]. It reflects the hydraulic feature of high capacitance. However, longer-term fluctuations in water status can lead to radial fluctuations in the sectional dimensions of tracheids [78]. A marked, brief decline in those dimensions represents a well-defined false ring. Similarly, the maritime pine, Pinus pinaster Aiton, has shown very marked within-ring fluctuations in tracheid dimensions under manipulated drought status [79]. In neither maritime pine nor P. radiata, however, did drought per se appear to have induced a negative phenological state.
3.10. Compression Wood and Feedback Effects of Ontogenetic Phenology
Like almost all conifers, P. radiata readily forms compression wood, especially with high soil fertility [62]. The compression wood [80], in mild cases, is more readily visible in LW than in EW. In severe cases, however, EW–LW differentiation is not visually clear. Also, Burdon [62] observed elevated compression wood incidence near ground level, suggesting that the juvenile growth habit is associated with increased propensity to form compression wood in P. radiata. The juvenile habit, which makes shoots less attractive to browsing mammals, also makes material more prone to bending and lodging, as in cereals. This is believed to contribute to vulnerability to snow damage, which would account for relatively poor performance on some snow-prone sites of material with persistent juvenility [81]. Any advantage in self-righting ability that such material might have was clearly not decisive. However, no attempt appears to have been made to exploit the postulated superior snow tolerance of cuttings with some degree of maturation.
4. Other Tree Species
Annual ring profiles, in terms of radial within-ring variations in xylem cell properties in different positions from the pith and from ground level, not only reflect phenology but also provide an informative basis for comparing the influences of phenology on within-ring variation in the properties among different species. In conifers, most wood properties are completely dominated by those of tracheids; the remaining cell types, in rays and resin canals representing very small proportions of the xylem. In hardwoods, the variations involve a wider range of cell types, notably the vessels and xylem fibres. I/we now consider some examples in categories of trees to illustrate the range of situations. Systematic review is not attempted.
The species considered below are admittedly a fragmentary and unrepresentative sample of tree taxa. Nevertheless, they illustrate the wide ranges of shoot-extension architecture and phenology and of within-ring wood variation in relation to the phenology. They also indicate the limitations of inferences based only on P. radiata and provide a basis for some broader, if tentative, conclusions.
The link between the phenology of shoot extension with the behaviour of the cambium is complicated by the variation among woody plant taxa in the architecture of the extending shoot extremities. It is clear, though, that whereas the degree of EW–LW differentiation tends to be associated with the seasonality of shoot extension, the association is very incomplete.
Also not addressed is the seasonality of accretion of extractives in heartwood formation and enrichment. Nor is the contribution of bark production to over-bark stem increment. Heartwood extractives in at least many cases confer resistance to stem decay. Thickened bark typically defends trees against fire and/or animal damage, e.g., [82]. As such, both features represent defences that presumably represent some diversion of resources from xylem production (cf [83]).
4.1. Other Conifers
Members of the Pinaceae generally have pronounced seasonal phenology, in the sense of producing sealed buds that burst in spring or early summer to produce flushes of shoot and foliage elongation. This is typically associated with pronounced EW–LW differentiation, although the contrasts involved are variable [5]. In the one deciduous genus, Larix, the contrast is predictably marked. In Douglas-fir, that differentiation is particularly marked (Figures 6–11 in [4,5]), The consequent “lamination” of alternating ring layers with contrasting properties may contribute to the noteworthy stiffness of its wood. Speculatively, the strong within-ring heterogeneity of Douglas-fir wood properties may help reconcile the competing requirements for capacitance, hydraulic conductivity, and resistance to embolism in a species that can grow to extreme heights (cf [30]). Yet coast redwood (Sequoia sempervirens (D.Don) Endl.), which can grow even taller, evidently has markedly less EW–LW contrast.
In the Cupressaceae/Taxodiaceae complex, the Cupressaceae (Cupressus and closely related genera) typically lack sealed buds, although seasonal growth flushes occur during which currently extending shoots are soft and unlignified. Despite typically less overt shoot elongation phenology, EW–LW differentiation is marked in species studied [5]. Additionally, as in Pinaceae, the degree of EW–LW differentiation differs widely [5] among such cases of similar shoot phenology. Alaska yellow-cedar (Chamaecyparis nootkatensis D.Don ex Lambert) shows much less within-ring density contrast than western redcedar (Thuja plicata Donn ex D.Don). For the Chilean conifer, alerce (Fitzroya cupressoides M.W., Johnst.), although densitometric data have not been located, the EW–LW differentiation is clearly very strong. In New Zealand Libocedrus, Patel [84] observed clear EW–LW differentiation. Among the Taxodiaceae, the dawn redwood (Metasequoia glyptostroboides Miki) has shown relatively low EW–LW density difference [5], despite a strong phenological manifestation in being deciduous.
4.2. Podocarpaceae
Among other gymnosperm families, the Podocarpaceae show variability in shoot extension architecture and in shoot phenology. This is illustrated by some New Zealand species. For these, annual ring density profiles have not been located, but photomicrographs of annual ring boundaries have been published (mainly Patel [85]), which allow rough evaluation of EW–LW differentiation. Rimu (Dacrydium cupressinum Sol. Ex Lamb.) and kahikatea (Dacrycarpus dacrydioides (A, Rich.) De Lamb.) do not produce sealed buds. For rimu, the annual rings are visually discernible and the LW is microscopically evident in branches [86]. In kahikatea, there is a progression to a narrow zone of moderately defined LW with a clear boundary with the next ring’s EW. By contrast, totara (Podocarpus totara G. Donn. ex D.Don) shows clearly episodic shoot extension interspersed with bud formation and quite marked EW–LW differentiation. However, matai (Prumnopitys taxifolia (Banks and Sol. ex D. Don) de Laub.) (syn, Podocarpus spicatus), which shows vestigial bud structures, showed minimal EW–LW differentiation.
4.3. Deciduous Hardwoods
Such trees in temperate zones form sealed buds and then shed foliage in response to declining temperatures and/or shortening days during autumn [12,14]. In spring, bud break may or may not require a period of winter chilling or vernalisation. Either way, it typically occurs after a period of sufficient temperatures, which are often characterised as “degree days”. In the tropics, the onset of seasonal drought can evidently provide the cue for bud set and foliage shedding, e.g., [87]. However, some species do the opposite, producing fresh foliage upon the onset of a dry season and even shedding their foliage during a wet season; for such behaviour, Burdon and Bartlett [9] have postulated pathogen avoidance. Yet, despite likely differences in environmental cues, tropical and temperate species overlap strongly in their within-ring variations in wood anatomy.
Some taxa, notably oaks (Quercus spp,), ashes (Fraxinus spp.), sweet chestnuts (Castanea spp.), which are almost all temperate, and teak (Tectona grandis L.), which is tropical, are ring porous, with a quite uniformly narrow zone of EW in which vessels predominate [5]. The LW, in which xylem fibres predominate, is typically more variable in its width and this tends to reflect the tree’s vigour. The EW and LW are formed during and after the initial spring flush, respectively. This means that wood density tends to be positively associated with ring width. Despite the anatomical differences within the ring, the EW–LW density contrast tends to be less than in conifers [5]. Among other deciduous hardwoods with similar shoot-extension phenology, there are genera that are diffuse porous [5]. These include birches (Betula spp.), alders (Alnus spp.), and maples (Acer spp.). In them, density varies little within rings, although growth rings may be visually discernible. Thus, their within-ring density profiles and whole-ring densities are much less governed by seasonal shoot phenology.
Where EW–LW differentiation is weak in respect of density, it mighty not necessarily be so for some other, less easily determined wood properties.
4.4. Evergreen Hardwoods
In this category, seasonal vegetative phenology in shoot extension is often weakly expressed, in there being no distinctive dormant bud structures. This situation is common among eucalypts (Eucalyptus app, Corymbia spp., Angophora spp.). Correspondingly, such species tend to have diffuse porous or semi-ring porous wood. For Eucalyptus muelleriana A.W.Howitt, Harris [29], using densitometry, observed no clear EW–LW differentiation. However, there are some evergreen hardwoods with strongly seasonal shoot–tip phenology. For two New Zealand species, red beech (Nothofagus fusca (Hook.f.)Oerst.) and silver beech (N. menziesii Hook.f.), which are diffuse-porous to semi-ring-pororus, photomicrogaphic evidence is available [88]. In both, the ring begins with a narrow concentration of relatively large vessels, followed by progressively smaller vessels, indicating a progressive increase in density until the boundary with the following ring, the progression being stronger in N. fusca. Thus EW–LW differentiation is weak.
5. An Overarching Question for Regulation of Diameter Growth
The most deeply intrinsic regulation of diameter growth involves the fact that in nature a tree must be a successful competitor. For competitive ability, many contributing factors have been identified as contributing to competitive ability [89]. In this context, however, the key factors are suspension of the tree crown so as to capture light in the presence of other trees and having the mechanical robustness to cope with the abiotic mechanical stresses occurring in the environment. The combined evolutionary pressures involved are proposed as the basis for what Mattheck [90] termed the “compromise tree”. That can typically entail producing the minimum of wood needed to securely suspend the tree crown in a competitive position, thereby maximising the mechanical efficiency of the stem [34,90]. Yet that mechanical efficiency is often achieved by variations in wood properties within the stem that are very troublesome for wood processing and utilisation. These variations involve, inter alia, wood density and its component traits and patchy occurrence of reaction wood with its anomalous mechanical and shrinkage properties. Classic examples within P. radiata are the corewood–outerwood progression in fibre properties and the formation of reaction wood (in this case compression wood). With the former, the outerwood “sheath” with high density and consequent stiffness and strength enclosing the intergrading corewood has evident mechanical efficiency, as traced theoretically by Lachenbruch et al. [30]. That is analogous, in crude terms, to a hollow pole being more mechanically efficient for a given amount of material than a solid one. With compression wood, it evidently serves a mechanical purpose, and its production can be an active “righting” mechanism for the stem [62,66]. For trees to be a truly efficient “wood factory”, they would need paradoxically to be mechanically inefficient, without the inconvenient within-stem variations in wood properties and producing wood “gratuitously”, in excess of mechanical requirements (Section 9.2.1 in [34]). That could prospectively eliminate or reduce unwanted variations in wood properties. It should also boost the harvest index, namely the usable proportion of primary biological production, assuming that the photosynthetic capacity of a healthy foliage canopy is largely fixed. Achieving all that would entail an obvious sacrifice of competitive ability but could be overcome by appropriate genetic deployment that averts inter-genotype crown competition. The simplest such deployment system would be growing trees not in monoclonal blocks but in principle mixtures of “silviculturally compatible” clones. Achieving crop efficiency at the expense of competitive ability is actually inherent to much traditional crop breeding, an extreme case being the dwarf cereals of the “Green Revolution” that is outlined by Gaud [91]. A big challenge for achieving such a goal with trees would lie in retaining some capability for self-righting in stems. That aside, it is difficult to see how phenology per se might play any substantial role in the feedback mechanisms whereby a species allocates resources in its strategy of balancing the needs of competitive ability and mechanical stability.
Another aspect of maintaining mechanical stability is the response to mechanical stresses/loads, which are known to stimulate diameter growth. In the field, guying experiments, reducing wind sway, have reduced stem diameter growth, e.g., [92,93]. Conversely, Cown [94] reported circumferential compression wood produced in P. radiata after thinning, attributed to increased wind sway, with a temporary boost in butt-log diameter growth. Despite the putative role of wind sway, flexural wood as defined by Telewski [95] was evidently not involved. In conifers, the compression wood shows reduced stiffness [66], though yielding more to tensile forces. In hardwoods, however, the reverse is the case with the tension wood, although not all species produce it [66].
Also studied has been the behaviour of xylem growth in correcting aberrant situations, such as correcting or preventing severe disorientation of stems or structural accommodation of physical obstructions, rot-created hollows or other physical damage [90].
There are mechanisms for trees to achieve mechanical stability other than purely through stem diameter growth in response to stresses, within-ring and pith-to-bark variation, and reaction wood or flexure wood. Growth stresses are one, they serve well the intact stem [90] but they can be disastrous for producing sawn timber. Flexibility of tree crowns can reduce wind stresses by reducing wind resistance, e.g., [96]. Also, I postulate that brittleness of branch bases, as in Douglas-fir, can serve to protect the rest of the tree by some of them snapping off and thereby reducing wind resistance.
By and large, these crown-property mechanisms for conferring mechanical resilience seem to be essentially independent of seasonal phenology. Ontogenetic phenology, however, appears to play some role. For instance, in the juvenile phase P. radiata appears to be more resilient to toppling, although more susceptible in the first instance, and to be both less palatable and more resilient to browsing [34,59].
Of great research interest, with potentially great economic payoffs, is elucidating the mechanisms whereby trees naturally balance the needs for competitive ability and mechanical stability. Knowledge of the mechanisms is likely to be crucial to changing the balance so as to substantially raise the level of domestication, which is still low in forest trees despite what tree breeders have achieved. Acquiring that knowledge poses a huge challenge, surely requiring a formidable amount of genomic and metabolomic research. Translating the requisite knowledge into quick, effective selection will presumably require much genomic information. Indeed, such information would likely help with elucidating the feedback mechanisms.
6. Conclusions
With the range of seasonal climatic fluctuations, the diversity of shoot and inflorescence architecture, and the range of biotic influences, the phenology of different tree species involves a huge range of adaptive strategies. Comparison among such strategies, however, is complicated by the fact that an adaptive strategy is never defined by individual features but rather by the interplay between the various features to constitute a valid and robust strategy.
The role of phenology per se, in governing radial growth, is seen as influencing within-ring and pith-to-bark variation in wood properties (much more in some species than in others), rather than governing amount of radial growth.
Although various physical, anatomical, and even chemical properties show within-ring variation, wood density is often particularly informative concerning the impacts of seasonal phenology.
The specific influence of the state of shoot phenology on wood properties, from pith to bark and within rings, although showing general trends, is variable even among related species.
In hardwoods, the behaviour of wood properties in relation to phenology seems more variable than in conifers, in keeping with their more complex and variable wood anatomy; however, the within-ring variation in density tends to be less.
How the shoot phenology of a species influences within-tree variation in wood properties appears to reconcile the phenological adaptation to climatic and biotic environments with the mechanical and hydraulic needs of the species.
Competitive requirements in trees are met by parsimony of wood production that is generally dominated by diameter growth. This parsimony often entails troublesome within-tree variation in wood properties. The need for competitive ability can be circumvented by clonal culture systems. However, intensifying domestication by making trees more efficient “wood factories” without unwanted within-tree variation, poses great challenges. In meeting them, however, major phenological constraints are not foreseen.
Funding
The work was supported by Scion providing office facilities to the author in his capacity as Emeritus Scientist. Scion also met the Article Processing Charge.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Thanks are due to Marlene Joubert for uploading photographic images, Dale Corbett for the graphics, and Elizabeth Miller for providing items of information.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Burdon, R.D.; Moore, J.R. Adverse Genetic Correlations and Impacts of Silviculture Involving Wood Properties: Analysis of Issues for Radiata Pine. Forests 2018, 9, 308. [Google Scholar] [CrossRef] [Green Version]
- Park, J.S.; Post, E. Seasonal timing on a cyclical Earth: Towards a theoretical framework for the evolution of phenology. PLoS Biol. 2022, 20, e3001952. [Google Scholar] [CrossRef] [PubMed]
- Denne, M.P. Definition of latewood according to Mork (1928). IAWA J. 1989, 10, 59–62. [Google Scholar] [CrossRef]
- Harris, J.M.; Cown, D.J. Basic wood properties. In Properties and Uses of New Zealand Radiata Pine. Vol1—Wood Properties; Kininmonth, J.A., Whitehouse, L.J., Eds.; New Zealand Ministry of Forestry: Wellington, New Zealand, 1991; pp. 6-1–6-28. [Google Scholar]
- Cown, D.J.; Parker, M.L. Comparison of annual ring density profiles in hardwoods and softwoods by X-ray densitometry. Can. J. For. Res. 1978, 8, 442–449. [Google Scholar] [CrossRef]
- Evans, R.; Downes, G. Recent developments in automated wood quality assessment. In Eucalypt Plantations and Improving Fibre Yield and Wood Quality, Proceedings of the IUFRO Conference, Hobart, Australia, 19–24 February 1995; Potts, B.M., Borralho, N.M.G., Reid, J.B., Cromer, R.N., Tibbits, W.N., Raymond, C.A., Eds.; Centre for Co-operative Research on Temperate Hardwood Forestry: Sandy Bay, Tasmania, Australia, 1995; pp. 101–105. [Google Scholar]
- Eberhardt, T.L.; Samuelson, L.J. Collection of wood quality data by X-ray densitometry: A case study with three southern pines. Wood Sci. Technol. 2015, 49, 739–753. [Google Scholar] [CrossRef]
- Burdon, R.D. Phenology of Pinus radiata—A Big Anomaly? Overview with Some Research Agenda. 2020. Available online: https://www.researchgate.net/publication/359222427_Phenology_of_Pinus_radiata_2022 (accessed on 7 March 2023).
- Burdon, R.D.; Bartlett, M.J. Putative biotic drivers of plant phenology: With special reference to pathogens and deciduousness. Ecol. Evol. 2022, 12, e8932. [Google Scholar] [CrossRef]
- Aide, T.M. Dry season leaf production: An escape from leaf herbivory. Biotropica 1994, 24, 532–537. [Google Scholar] [CrossRef]
- Lamarre, G.P.A.; Mondoza, I.; Fine, P.V.A.; Baraloto, C. Leaf synchrony and insect herbivory among tropical tree specialists. Plant Ecol. 2014, 215, 208–220. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S.G. Growth Control in Woody Plants; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Wareing, P.F. Photoperiodism in woody plants. Annu. Rev. Plant Physiol. 1956, 7, 191–214. [Google Scholar] [CrossRef]
- Thomas, B.; Vince-Prue, D. Photoperiodism in Plants, 2nd ed.; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Fordham, R.A.; Ogden, J. An Ecological Approach to New Zealand’s Future; New Zealand Ecological Society: Christchurch, New Zealand, 1974; Volume 21. [Google Scholar]
- Borchert, R.; Renner, S.S.; Calle, Z.; Navarrete, D.; Tye, A.; Gautier, L.; Spichiger, R.; von Hildebrand, P. Photoperiodic induction of synchronous flowering near the Equator. Nature 2005, 433, 627–629. [Google Scholar] [CrossRef]
- Calle, Z.; Schlumpberger, B.O.; Piedrahita, L.; Leftin, A.; Hammer, S.A.; Borchert, R. Seasonal variation in daily insolation induces synchronous bud break and flowering in the tropics. Trees 2010, 24, 865–877. [Google Scholar] [CrossRef]
- Borchert, R.; Calle, Z.; Strahler, A.H.; Baertschi, A.; Magill, R.E.; Broadhead, J.S.; Kamau, J.; Nioroge, J.; Muthuri, C. Insolation and photoperiodic control of tree development near the equator. New Phytol. 2015, 205, 7–13. [Google Scholar] [CrossRef]
- Wareing, P.F. Tree physiology in relation to genetics and breeding. Unasylva 1964, 18, 61–76. [Google Scholar]
- Poethig, R.S. Phase change and the regulation of morphogenesis in plants. Science 1990, 250, 923–930. [Google Scholar] [CrossRef]
- Greenwood, R.M. Some differences between plants of the Chatham Islands and the New Zealand mainland. N. Z. J. Ecol. 1992, 16, 51–52. [Google Scholar]
- Burns, K.C.; Dawson, J.W. Heteroblasty on Chatham Island: A comparison with New Zealand and New Caledonia. N. Z. J. Ecol. 2009, 33, 156–163. [Google Scholar]
- Norskov-Lauritsen, G. Notes on the growth habits of the main pine species of the Eastern Transvaal Region. For. S. Afr. 1963, 3, 67–83. [Google Scholar]
- Larson, P.R. A physiological consideration of the springwood summerwood transition in Pinus resinosa. For. Sci. 1960, 6, 110–122. [Google Scholar]
- Larson, P.R. Wood Formation and the Concept of Wood Quality; Bulletin 74; Yale University School of Forestry: New Haven, CT, USA, 1969. [Google Scholar]
- Larson, P.R. The Vascular Cambium: Development and Structure; Springer: Berlin/Heidelberg, Germany, 1994; pp. x11 + 723. [Google Scholar]
- Little, C.H.A.; Savidge, R.A. The role of plant growth regulators in forest tree cambial growth. In Hormonal Control of Tree Growth; Kossuth, S.V., Ross, S.D., Eds.; Forestry Sciences Series; Springer: Berlin/Heidelberg, Germany, 1987; Volume 28, pp. 137–169. [Google Scholar]
- Savidge, R.A. Intrinsic regulation of diameter growth. J. Plant Growth Regul. 2001, 29, 52–77. [Google Scholar] [CrossRef]
- Harris, J.M. The use of beta rays in determining wood properties—Part 2. Measuring earlywood and latewood. N. Z. J. Sci. 1969, 12, 409–418. [Google Scholar]
- Lachenbruch, B.; Moore, J.R.; Evans, R. Radial variation in wood structure and function in woody plants, and hypotheses for its occurrence. In Size- and Age-Related Changes in Tree Structure and Function; Meinzer, F.C., Lachenbruch, B., Dawson, T.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 121–164. [Google Scholar]
- Scott, C.W. Pinus radiata; FAO: Rome, Italy, 1960. [Google Scholar]
- Burdon, R.D. Pinus radiata. In Ecosystems of the World, Tree Crop Ecosystems; Last, F.T., Ed.; Elsevier: Amsterdam, The Netherlands, 2001; Volume 19, pp. 99–161. [Google Scholar]
- Mead, D.J. Sustainable Management of Pinus radiata Plantations; FAO Forestry Paper 170; FAO: Rome, Italy, 2013. [Google Scholar]
- Burdon, R.D.; Libby, W.J.; Brown, A.G. Domestication of Radiata Pine; Springer International: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- FAO. FAOSTAT FAO Forestry Production and Trade Statistics; FAO: Rome, Italy, 2018. [Google Scholar]
- Burdon, R.D. Genetic survey of Pinus radiata. 9. General discussion and implications for genetic management. N. Z. J. For. Sci. 1992, 22, 274–298. [Google Scholar]
- Madgwick, H.A.I. Pinus radiata—Biomass, Form and Growth; H.A.I. Madgwick: Rotorua, New Zealand, 1994. [Google Scholar]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. Hortic. Sci. 1987, 22, 371–377. [Google Scholar]
- Jewaria, P.K.; Hanninen, H.; Li, X.; Bhalerao, R.P.; Zhang, R. A hundred years after: Endodormancy and the chilling requirement in tropical trees. New Phytol. 2021, 231, 565–570. [Google Scholar] [CrossRef] [PubMed]
- MacDougal, D.T. Tree Growth; Chronica Botanica: Leiden, The Netherlands, 1938. [Google Scholar]
- Fielding, J.M.; Millett, M.R.O. Some Studies of the Growth of Monterey Pine. Part 1. Diameter Growth; Bulletin 27; Commonwealth Forestry Bureau: Canberra, Australia, 1941. [Google Scholar]
- Fielding, J.M. The Seasonal and Daily Elongation of the Shoots of Monterey Pine and the Daily Elongation of the Roots; Leaflet No. 75; Forestry and Timber Bureau: Canberra, Australia, 1955. [Google Scholar]
- Pawsey, C.K. Height and diameter growth cycles in Pinus radiata. Aust. For. Res. 1964, 1, 3–8. [Google Scholar]
- Cremer, K.W. Seasonal variation of shoot developments in Pinus radiata near Canberra. Aust. For. Res. 1973, 6, 31–52. [Google Scholar]
- Bollmann, M.P.; Sweet, G.B. Bud morphogenesis of Pinus radiata in New Zealand. II. The seasonal shoot growth pattern of seven clones at four sites. N. Z. J. For. Sci. 1979, 9, 153–165. [Google Scholar]
- Bollmann, M.P.; Sweet, G.B.; Rook, D.A.; Halligan, E.A. The influence of temperature, nutrient status, and short drought on seasonal initiation of primordia and shoot elongation in Pinus radiata. Can. J. For. Res. 1986, 16, 1019–1029. [Google Scholar] [CrossRef]
- Burdon, R.D. Annual growth stages for height and diameter in Pinus radiata. N. Z. J. For. Sci. 1994, 24, 11–17. [Google Scholar]
- Jacobs, M.R. The Detection of Annual Growth Stages in the Crown of Pinus radiata; Bulletin No. 19; Commonwealth Forestry Bureau: Canberra, Australia, 1937. [Google Scholar]
- Bannister, M.H. Some variations in the growth pattern of Pinus radiata in New Zealand. N. Z. J. Sci. 1962, 5, 342–370. [Google Scholar]
- Kummerow, J. Growth anomalies of Pinus radiata under tropical conditions. Proc. Ger. Bot. Soc. 1962, 75, 37–40. [Google Scholar]
- Lanner, R.M. The phenology and growth habits of pines in Hawaii; Research Paper PSW-31; USDA Forest Service: Washington, DC, USA, 1966. [Google Scholar]
- Barnett, J.R. Winter activity in the cambium of Pinus radiata. N. Z. J. For. Sci. 1971, 1, 208–222. [Google Scholar]
- Bamber, R.K.; Burley, J. The Wood Properties of Radiata Pine; Commonwealth Agriculture Bureaux: Slough, UK, 1983; pp. vi + 84. [Google Scholar]
- Nanayakkara, B.; Dickson, A.R.; Meason, D.F. Xylogenesis of Pinus radiata D. Don growing in New Zealand. Ann. For. Sci. 2019, 76, 74. [Google Scholar] [CrossRef]
- Harris, J.M. A Survey of Wood Density, Tracheid Length, and Latewood Characteristics of Radiata Pine Grown in New Zealand; FRI Technical Paper 47; New Zealand Forest Service: Wellington, New Zealand, 1965. [Google Scholar]
- Cown, D.J.; McConchie, D.L.; Young, G.D. Radiata Pine Wood Properties Survey; FRI Bulletin; New Zealand Ministry of Forestry: Wellington, New Zealand, 1991; Volume 50. [Google Scholar]
- Harris, J.M. The use of beta rays in determining wood properties—Part 3. Summarising radial variations in wood density. N. Z. J. Sci. 1969, 12, 419–431. [Google Scholar]
- Burdon, R.D.; Bannister, M.H.; Low, C.B. Genetic survey of Pinus radiata. 2. Population comparisons for growth, disease resistance, and morphology. N. Z. J. For. Sci. 1992, 22, 119–137. [Google Scholar]
- Burdon, R.D.; Bannister, M.H.; Low, C.B. Genetic survey of Pinus radiata. 3. Variance structures and narrow-sense heritabilities for growth variables and morphological traits in seedlings. N. Z. J. For. Sci. 1992, 22, 160–186. [Google Scholar]
- Burdon, R.D.; Bannister, M.H.; Low, C.B. Genetic survey of Pinus radiata. 4. Variance structures and heritabilities in juvenile clones. N. Z. J. For. Sci. 1992, 22, 187–211. [Google Scholar]
- Burdon, R.D.; Kibblewhite, R.P.; Walker, J.C.F.; Megraw, R.; Cown, D.J. Juvenile versus mature wood: A new concept, with special reference to Pinus radiata and P. taeda. For. Sci. 2004, 50, 399–415. [Google Scholar]
- Burdon, R.D. Compression wood in Pinus radiata clones on four different sites. N. Z. J. For. Sci. 1975, 5, 152–164. [Google Scholar]
- Will, G.M.; Hodgkiss, P.D. Influence of nitrogen and phosphorus on growth and form of radiata pine. N. Z. J. For. Sci. 1977, 7, 307–320. [Google Scholar]
- McConchie, D.L. Wood Quality of Radiata Pine Grown on Farm Sites; Klitscher, K., Cown, D., Donaldson, L., Eds.; FRI Bulletin 201; Wood Quality Workshop: Rotorua, New Zealand, 1996; pp. 38–45. [Google Scholar]
- Burdon, R.D. Wood properties and genetic improvement of radiata pine. N. Z. J. For. 2010, 55, 22–27. [Google Scholar]
- Donaldson, L.A.; Singh, A.P. Reaction wood. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 93–110. [Google Scholar]
- Greer, D.H. Temperature regulation of the development of frost hardiness in Pinus radiata D. Don. Aust. J. Plant Physiol. 1983, 10, 539–547. [Google Scholar] [CrossRef]
- Greer, D.H.; Warrington, I.J. Effect of photoperiod, night temperature, and frost incidence on development of frost hardiness in Pinus radiata. Aust. J. Plant Physiol. 1982, 9, 333–342. [Google Scholar] [CrossRef]
- Greer, D.H.; Stanley, C.J.; Warrington, I.J. Photoperiodic control of the initial phase of frost hardiness development in Pinus radiata. Plant Cell Environ. 1989, 12, 661–668. [Google Scholar] [CrossRef]
- Menzies, M.I.; Holden, D.G. Seasonal frost-tolerance of Pinus radiata, Pinus muricata, and Pseudotsuga menziesii. N. Z. J. For. Sci. 1981, 11, 82–99. [Google Scholar]
- Jenkins, P.A.; Hellmers, H.; Edge, E.A.; Rook, D.A.; Burdon, R.D. Influence of photoperiod on growth and wood formation in Pinus radiata. N. Z. J. For. Sci. 1977, 7, 172–191. [Google Scholar]
- Burdon, R.D. Photoperiodic effect on pollen shedding in Pinus radiata? N. Z. J. For. Sci. 1977, 7, 214–215. [Google Scholar]
- Schimleck, L.R.; Dahlen, J.; Auty, D. Radial patterns of specific gravity variation in North American conifers. Can. J. For. Res. 2022, 52, 889–900. [Google Scholar] [CrossRef]
- Cown, D.J. Variation in tracheid dimensions in the stem of a 26-year-old radiata pine tree. Appita 1975, 28, 237–245. [Google Scholar]
- Drew, D.M.; Downes, G. A model of stem growth and wood formation in Pinus radiata. Trees 2015, 29, 1395–1413. [Google Scholar] [CrossRef]
- Jackson, D.S.; Gifford, H.H.; Chittenden, J. Environmental factors influencing the increment of Pinus radiata (2) Effects of seasonal drought on height and diameter increment. N. Z. J. For. Sci. 1976, 6, 265–286. [Google Scholar]
- Rook, D.A.; Swanson, R.H.; Cranswick, A.M. Reaction of radiata pine to drought. Inf. Ser. N. Z. Dep. Sci. Ind. Res. 1977, 126, 55–68. [Google Scholar]
- Downes, G.M.; Evans, R. Effects of environment on tracheid dimensions. In Proceedings of the Workshop ’Modelling of Tree-ring Development—Cell Structure and Environment’; Spieker, H., Kahle, P., Eds.; Institut für Waldwachstum: Freiberg, Germany, 1994. [Google Scholar]
- Vieira, J.; Moura, M.; Nabais, C.; Freitas, H.; Campelo, F. Seasonal adjustment of primary and secondary growth in maritime pine under simulated climatic changes. Ann. For. Sci. 2019, 76, 84. [Google Scholar] [CrossRef]
- Timell, T.E. Origin and Evolution of Compression Wood; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Burdon, R.D.; Firth, A.; Low, C.B.; Miller, M.A. Native provenances of Pinus radiata in New Zealand: Performance and potential. N. Z. J. For. 1997, 41, 32–36. [Google Scholar]
- Gumber, S.; Singh, R.D.; Ram, J.; Tewari, A.; Singh, S.P. Bark thickness analysis of four dominant tree species of Central Himalayan forests varying in exposure to surface fires. Trees 2022, 36, 685–695. [Google Scholar] [CrossRef]
- Loehle, C.; Namkoong, G. Constraints on tree breeding: Growth tradeoffs, growth strategies, and defensive investments. For. Sci. 1987, 33, 1089–1097. [Google Scholar]
- Patel, R.N. Wood anatomy of Cupressaceae and Araucariaceae indigenous to New Zealand. N. Z. J. Bot. 1968, 6, 9–18. [Google Scholar] [CrossRef]
- Patel, R.N. Wood anatomy of Podocarpaceae indigenous to New Zealand. N. Z. J. Bot. 1967, 5, 307–321. [Google Scholar] [CrossRef]
- Eom, Y.G.; Butterfield, B.G. Anatomical comparison of compression, opposite and lateral wood in New Zealand rimu (Dacrydium cupressinum Lamb.). Mokchae Konghak 2001, 29, 1–13. [Google Scholar]
- Ghosh, S.; Nandy, S.; Mohanti, S.; Subba, R.; Kushwaha, S.P.S. Are phenological variations in natural teak (Tectona grandis) forests of India governed by rainfall? Environ. Monit. Assess. 2020, 191 (Suppl. S3), 786. [Google Scholar] [CrossRef]
- Patel, R.N. Wood anatomy of the dicotyledons indigenous to New Zealand 15. Fagaceae. N. Z. J. Bot. 1986, 24, 189–202. [Google Scholar] [CrossRef]
- Simon, J.; Schmidt, S. Plant competition in a changing world. Front. Plant Sci. 2017, 8, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattheck, C. Trees: The Mechanical Design; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Gaud, W.S. The green revolution: Accomplishments and apprehensions. In AgrobioBioWorld; The Society for International Development: Washington, DC, USA, 1968. [Google Scholar]
- Jacobs, M.R. The effect of wind sway on the form and development of Pinus radiata D. Don. Aust. J. Bot. 1954, 2, 35–51. [Google Scholar] [CrossRef]
- Moore, J.R.; Cown, D.J.; Lee, J.R.; McKinley, R.B.; Brownlie, R.K.; Jones, T.G.; Downes, G.M. The influence of stem guying on radial growth, stem form and internal resin features in radiata pine. Trees 2014, 28, 1197–1207. [Google Scholar] [CrossRef]
- Cown, D.J. Comparison of the effects of two thinning regimes on some wood properties of radiata pine. N. Z. J. For. Sci. 1974, 4, 540–551. [Google Scholar]
- Telewski, F.W. Flexure wood: Mechanical stress induced secondary xylem formation. In Secondary Xylem Biology; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 73–91. [Google Scholar]
- Gardiner, B.; Berry, P.; Moulia, B. Wind impacts on plant growth. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef]
Figure 1.
Young stand of P. radiata being pruned, showing general tree form (Scion Image Library).

Figure 2.
Old stand of P. radiata (>40 years), showing general tree form (Scion Image Library).

Figure 3.
Young seedling of P. radiata, showing green apical tuft and a predominance of primary foliage in a neutral phenological state typical of Pinaceae (Scion Image Library).
Figure 3.
Young seedling of P. radiata, showing green apical tuft and a predominance of primary foliage in a neutral phenological state typical of Pinaceae (Scion Image Library).

Figure 4.
Graft of P. radiata, showing adult shoot morphology of the tree from which scion was taken, with entirely secondary foliage, sealed buds, and seed cones. Shoot growth has become quite strongly seasonal, but the top shows features of a “basal flush” (Scion Image Library).
Figure 4.
Graft of P. radiata, showing adult shoot morphology of the tree from which scion was taken, with entirely secondary foliage, sealed buds, and seed cones. Shoot growth has become quite strongly seasonal, but the top shows features of a “basal flush” (Scion Image Library).

Figure 5.
Graft of P. radiata, showing tip of leader during autumn with well-developed “basal flush” [44] subtending compound sealed bud containing lateral buds at tip (Scion Image Library).
Figure 5.
Graft of P. radiata, showing tip of leader during autumn with well-developed “basal flush” [44] subtending compound sealed bud containing lateral buds at tip (Scion Image Library).

Figure 6.
Densitometric traces showing within-ring progressions of wood density in (redrawn from Harris Figure 2 in [29]): Upper: P. radiata corewood, Lower: P. radiata outerwood.
Figure 6.
Densitometric traces showing within-ring progressions of wood density in (redrawn from Harris Figure 2 in [29]): Upper: P. radiata corewood, Lower: P. radiata outerwood.

Figure 7.
Pith-to-bark densiometric trace of P. radiata (redrawn from Bamber and Burley Figure 13 in [53]) showing both within-ring and corewood-to-outerwood trends.
Figure 7.
Pith-to-bark densiometric trace of P. radiata (redrawn from Bamber and Burley Figure 13 in [53]) showing both within-ring and corewood-to-outerwood trends.

Figure 8.
Stem disc of P. radiata, showing) increasing proportion of latewood in rings from pith to bark, and wide zone of sapwood, which confers high hydraulic capacitance. (Scion Image Library).
Figure 8.
Stem disc of P. radiata, showing) increasing proportion of latewood in rings from pith to bark, and wide zone of sapwood, which confers high hydraulic capacitance. (Scion Image Library).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
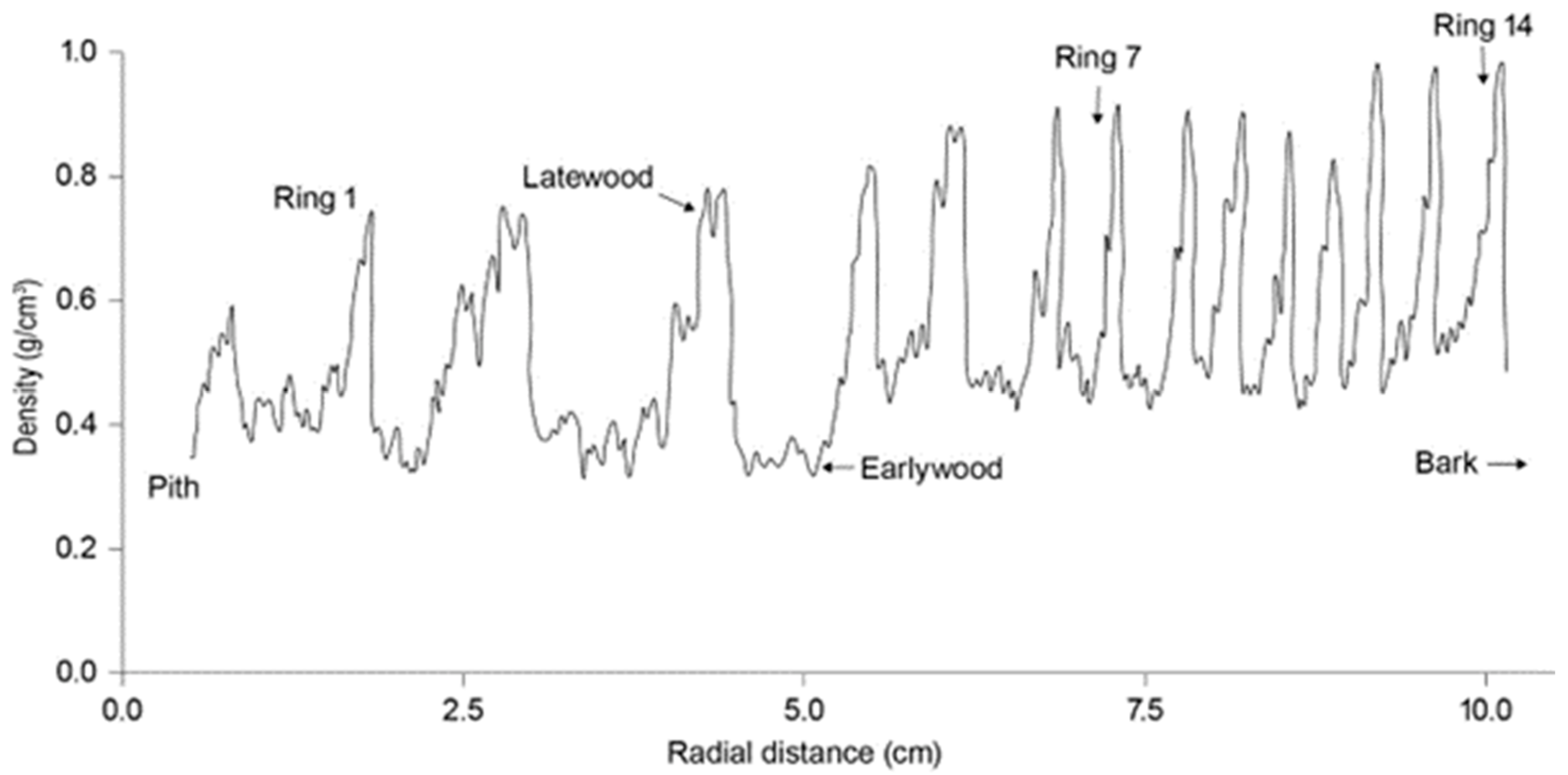{kind=link}
{kind=link}
Table 1.
Particulars of natural populations of P. radiata (from Burdon et al., [34]).
Table 1.
Particulars of natural populations of P. radiata (from Burdon et al., [34]).
| Population | Lat. (°N) | Elevation | Area (ha) | |
|---|---|---|---|---|
| (m a.s.l.) | Original | Current | ||
| Año Nuevo | 37 | 10–330 | 450 | 450 |
| Monterey | 361/2 | 10–580 | 3000 | 7400 |
| Cambria | 351/2 | 10–200 | 800 | 1400 |
| Guadalupe I. | 29 | 330–1155 | 640 | <200 trees * |
| Cedros I. | 28 | 275–640 | 130 | 130 |
* Now interspersed with young regeneration.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Burdon, R.D. Shoot Phenology as a Driver or Modulator of Stem Diameter Growth and Wood Properties, with Special Reference to Pinus radiata. Forests 2023, 14, 570. https://doi.org/10.3390/f14030570
AMA Style
Burdon RD. Shoot Phenology as a Driver or Modulator of Stem Diameter Growth and Wood Properties, with Special Reference to Pinus radiata. Forests. 2023; 14(3):570. https://doi.org/10.3390/f14030570
Chicago/Turabian StyleBurdon, Rowland D. 2023. "Shoot Phenology as a Driver or Modulator of Stem Diameter Growth and Wood Properties, with Special Reference to Pinus radiata" Forests 14, no. 3: 570. https://doi.org/10.3390/f14030570
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.